Research Article - (2024) Volume 7, Issue 2
Received: 01-Apr-2024, Manuscript No. jbr-24-132029;
Editor assigned: 03-Apr-2024, Pre QC No. P-132029;
Reviewed: 17-Apr-2024, QC No. Q-132029;
Revised: 22-Apr-2024, Manuscript No. R-132029;
Published:
29-Apr-2024
, DOI: 10.37421/2684-4583.2024.7.244
Citation: Titovets, Ernst. “A Novel Modell of Brain Cortex Water Metabolism Based on the Nanofluidic-domain Theory and its Biomedical Implications.’ J Brain Res 7 (2024): 244.
Copyright: © 2024 Titovets E. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Brain Water Metabolism (BWM) is involved in intercellular communication, transit of substrates, gases, drug delivery, spreading of malignant tumors. Various neurological conditions are accompanied by impaired BWM (e.g. idiopathic normal pressure hydrocephalus, syringomyelia, migraine, stroke, Alzheimer’s disease, etc.) The Extracellular Fluid (ECF), surrounding the neurocytes and the glia, circulates in the nanodimentional Extracellular Space (ECS), where its flow obeys the slip-flow principles of nanofluidics. The conventional view argues, however, that the nanodimentional ECS presents a diffusion barrier where convention is prohibited. A computer model has been developed revealing new features of BWM. The references are also presented, listing the AQP4-targeted drugs to be used in the therapy of brain edema. The research results may find their use in wide areas of neurobiological research, the drug therapy of water-metabolism-disorder conditions and targeted drug delivery.
Brain nanofluidic domain • Brain water metabolism • Brain water-metabolism-disorder conditions • Computational modelling • Targeted drug therapy
AQP4: Aquaporin-4; BWM: Brain Water Metabolism; CNM: Capillary Network Module; FS: Filtration Section; ISF: Interstitial Fluid; ISS: Interstitial Space; OS: Oscillatory Section; RS: Reabsorption Section
With its long research history, brain water metabolism still harbours some important unresolved issues. One of those is cantered on the events taking place in the brain extracellular space. In the words of Charles Nicholson the brain extracellular space is “the final frontier of neuroscience” [1]. It is an important admission on the part of this authority in neuroscience given with a suggestion of an adventure and discoveries associated in our minds with the last frontier.
Other researchers second this contention stating that: “Although neurons attract the most attention in neurobiology, our current knowledge of neural circuit can only partially explain the neurological and psychiatric conditions of the brain. Thus, it is also important to consider the influence of brain interstitial system, which refers to the space among neural cells and capillaries.” [2].
A stumbling block on the way of understanding functions of the nanodimentional Extracellular Space (ECS) is the state of water and its mobility there. The conventional bulk-water-approach theory asserts that water advection in this space asks for unrealistically high hydrostatic pressure gradients. Thus, fluid flow there is considered problematic and is even prohibitory on the thermodynamics grounds [3]. According to the conventional approach, the nanodimentional ECS presents a diffusion barrier where the narrowness of the spaces, with their characteristic width of 20–60 nm, and their tortuosity do not allow for any significant fluid flow [3-7].
On the other hand, nanofluidics, [8-10], a new discipline that came to fruition over the past two decades, suggests a different view on the events taking place in ECS. This rapidly developing science, dealing with fluid properties in the compartments with characteristic dimension 1 nm–100 nm, demonstrates that nan confinement is conducive to fast water flow. It is due to the slip-flow effect realized in such confinements [8]. Nanofluidics introduces new dictates for water dynamics in nanodimentional spaces. It gives grounds to consider ECS a nanofluidic domain where fluid flow is governed by the principles of nanofluidics [11-13].
This contention is supported by experimental results demonstrating circulation, in a pulsatile manner, of the Cerebrospinal Fluid (CSF) also involving Extracellular Fluid (ECF). There is observed continuous fluid exchange between the microvasculature, interstitial fluid and CSF [13,14]. An important role in brain water metabolism belongs to aquaporin AQP4. It ensures fast water exchange across the Blood-Brain Barrier (BBB) [15,16]. Aquaporins, the biological nanochannels, along with their artificial counterparts –nanotubes, have been widely used in nanofluidics research [17]. It is interesting to observe that, at one time, the biological channels were the channels to beat for the developers of their artificial counterparts. This ambitious task was eventually successfully accomplished with a development of even faster water-transferring artificial carbon-nanotube channels [18]. Computer simulation of brain water metabolism, conducted in this research, is based on the interdisciplinary approach and the theoretical principles of nanofluidics.
Experimental procedures
Theoretical assumptions and experiment-based parameters of the model:
- The brain interstitial extracellular space presents a nanofluidic domain where fluid movement is governed by the principles of nanofluidics.
- The hydrostatic pressure difference between the intracranial pressure ant that in the blood microvessels presents a driving force for isosmotic fluid exchange between the blood and ECS.
- Aquaporin AQP4 ensures kinetic control over water movement between the blood and the brain interstitial space.
- A phenomenological fluid flux equation, based on the Kedem–Katchalsky formalism of irreversible thermodynamics [19], has been used to describe water movement between the blood and ECS.
The quantitative data, describing geometric and thermodynamic parameters of the model, present the experimental observations available from the literature, as indicated further in this paper. A phenomenological flux equation describes water movement between the blood and ECS:
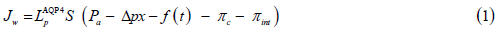
Where:
Jw – Water volumetric flow rate per unit transfer area, cm3⁄ s ⁄ mmHg;
AQP4 Lp – AQP4-dependent hydraulic conductivity coefficient
LpAQP4 = 13.7 × 10−6 for capillaries, 8.05 × 10-6 cm/s/mmHg for the arteriole and
LpAQP4 = 4.0 ×10−6 cm mmHg-1 s-1 for the draining venule);
S – Water transfer area, cm2,
Pa – Hydrostatic pressure at the arterial end of a microvessel, mmHg;
Δp – Hydrostatic pressure gradient along the longitudinal axis of a microvessel, mmHg/cm;
χ– The length of the longitudinal axis of a microvessel, cm;
πc – Capillary blood oncotic pressure, mmHg;
πint – interstitial fluid oncotic pressure, mmHg;
f(t) – The ICP hydrostatic pressure waveform (Figure 1)

Figure 1. A) Geometry of the capillary network module and B) The waveform of the hydrostatic intracranial pressure.
The overall water volume transferred over a period of one heart cycle is found upon numerical integration of the Eqn. 1 The model has been developed and numerically solved using the program Wolfram Mathematica 10.
We simulate water metabolism in a specified volume of the brain cortex that is viewed as a cuboid model. In the microvascular structure of this model, the capillary system is presented as a number of the Capillary Network Modules (CNM) in a respective cortex layer (Figure 1A and Figure 2). The microvascular geometry in CNM is modelled as a quasi-fractal tree stemming from the penetrating arteriole a (Figure 1A). This quasi-fractal array spreads on over sections S1-S5 of the module. Having mirrored further over the dashed line, it continues over sections S5-S9 to finally converge on the draining venule V. Detailed dimensions of the micro vessels in each section of CNM is given in Table 1. All micro vascular structures in the cuboid model are subjected to the oscillations of the intracranial hydrostatic pressure as shown in Figure 1B.

Figure 2. 3D brain cortex cuboid model with its vascularization network. The band arrows indicate the positions of the CNMs within their respective cortex layers (see Table 2 for details). Dimensions of the model: 0.030 × 0.030 × 0.125 cm and its volume is 11.2 × 10-5 cm3. The microvasculature occupies 3.26% of the model volume.
| Sections of the CNM | Capillary Radius, cm |
Capillary Length, cm |
The Number of the Capillaries in a Respective Section |
|---|---|---|---|
| S1 | 0.0009 | 0.00600 | 1 |
| S2 | 0.0005 | 0.00574 | 2 |
| S3 | 0.0004 | 0.00574 | 4 |
| S4 | 0.0004 | 0.00574 | 8 |
| S5 | 0.0004 | 0.01148 | 32 |
| S6 | 0.0004 | 0.00574 | 8 |
| S7 | 0.0004 | 0.00574 | 4 |
| S8 | 0.0005 | 0.00574 | 2 |
| S9 | 0.0009 | 0.00600 | 1 |
The peak of the wave, presented in Figure 1B, lies at 15 mmHg, while its base pressure is 3 mmHg. Duration of a complete heart cycle is 1.0 s. The waves of the intracranial hydrostatic pressure are generated by heart activity and closely follow the heart cycle phases [20]. Thus, the span over 0-0.143 s is related to the systolic phase and that over 0.143 s - 1.0 s –to the diastolic one. The wave was digitized to obtain its analytical presentation for further use in our simulations Table 1.
The cuboid model under consideration presents a construct of four penetrating arterioles, a central draining venule and a number of CNMs in a respective cortex layer (Figure 2). The band arrows indicate the positions of the CNMs within their respective cortex layers (see Table 2 for details). Dimensions of the model: 0.030 × 0.030 × 0.125 cm and its volume is 11.2 × 10-5 cm3. The microvasculature occupies 3.26% of the model volume (Figures 3 and 4).
| Brain Cortex Layers |
Thickness of the Layers x 10-1 cm |
Names of the CNM Placement Levels | Position of the Levels, cm |
Number of CNMs | Hydrostatic Pressure at Pa* mmHg | Hydrostatic Pressure at Pv** mmHg |
Hydrostatic Pressure Gradient along the Respective CNM, mmHg/cm |
|---|---|---|---|---|---|---|---|
| Layer I | 0.235 | Level-1 | 0.0118 | 2 | 60.5 | 8.4 | 900.5 |
| Layer II | 0.295 | Level-2 | 0.0375 | 4 | 51 | 8.6 | 732.3` |
| Level-3 | 0.0493 | 4 | 46 | 8.0 | 656.3 | ||
| Layer IIIa | 0.405 | Level-4 | 0.0938 | 3 | 30 | 9.5 | 354.1 |
| Level-5 | 0.1062 | 3 | 25 | 9.7 | 264.2 | ||
| Layer IIIb | 0.370 | Level-6 | 0.1250 | 4 | 18 | 10.0 | 138.2 |

Figure 3. 3D presentation of water exchange between brain cortex microvessels and ECS. A) Water exchange between the penetrating arteriole and ECS. The infinite planes p2, p3 delineate the Oscillatory Section (OS). V) Water exchange between the draining venule and ECS. The infinite plane indicated by p1 divides the systolic and the diastolic phases.

Figure 4. A top view of the 3D graphs in Figure 3 demonstrating topography of water filtration and reabsorption over the surface of the A) Penetraining arteriole and V) The draning venule. FS – Filtration Section; OS – Oscillatory Section; RS – Reabsorption Section. p1 indicates position of the infinite plane dividing the Systolic (S) and the Diastolic (D) phases. Water filtration –  ; Water reabsorption –
; Water reabsorption –
The thickness of the layers was selected after the dimension of those in the human brain neocortex [19]. The diameter of the four penetrating arterioles was taken to be 20 μm, remaining constant over the length of the micro vessel. The diameter of the draining venule was 40 μm. The simulated microvascular parameters, a layout of the capillaries, their distribution and density were essentially selected in keeping with the experimental observations [20,21]. In this microvascular arrangement that penetrating arterioles present a limiting step in the supply of blood to neocortex [19]. The simulation results in Figure 5 make it possible to see in some detail the character of water exchange with oscillatory section of the penetrating arteriole. As has been suggested earlier [22], wihin the oscillatory section there takes place a fast information exchange between the blood and the brain parenchyma.

Figure 5. Water movement in the oscillatory section of the penetrating arteriole. A) 3D presentation of water exchange and B) Top view of the 3D graph. The infinite plane p1 divides the systolic and the diastolic phases. Filtration - ; Reabsorption -
; Reabsorption -
As seen in Figure 6 the Oscillatory Section (OS) tends to migrate towards the arterial side of the capillary network with each deeper layer. Physiological significance of this OS shift is still to be elucidated. The results presented in Figure 6 and Figure 7 demonstrates that, within the top layers of the brain cortex, the capillary networks mostly participate in water filtrations. In the deeper cortex layers the process is reversed with the microvasculature absorbing outside water.

Figure 6. Capillary water metabolism at various levels of the brain cortex. The simulation results of water metabolism at the capillary networks in various cortex layers are presented as projections of their respective 3D graphs on their xoy plains (the top views of the 3D graphs). Filtration – ; Reabsorption –

Figure 7. Water metabolism at the capillary network of the various brain cortex layers. Water filtration - ; Water reabsorption -
; Water reabsorption -
The results in Figure 8 demonstrate that, within the selected geometry of the brain cortex and the thermodynamic parameters, there is generated an integral positive flux of ECF. This speaks against the classical dogma of the choroid plexus being the primary cerebrospinal fluid secretor.

Figure 8. Overall water fluxes in the cuboid model of the brain cortex.
a) Positive flux (water filtration) is 8.81 × 10-8 cm3/s; b) Negative flux (water reabsorption) is -1.01 × 10-8 cm3/s and c) Net positive flux is 7.72 × 10-8cm3/s.
As was outlined earlier in the Introduction section, the brain extracellular space, “the final frontier of neuroscience” [1], suddenly became a debatable issue as far as its function, physiological and pathological significance were concerned [23]. According to the conventional theory, the unidirectional CSF flow starts from the choroid plexus, that serves as CSF primary secretor, while the arachnoid granulations function as a primary collective CSF reabsorption site, channelling water into the blood stream [12,24]. This mechanism is central to the conventional theory of brain water metabolism.
Available now multiple experimental results demonstrate that the choroid plexus does not play any significant role in CSF circulation [25]. Large water fluxes continuously circulate in the whole of brain parenchyma. This water exchange involves all water-containing compartments of the brain including the blood microvessels and CSF [24]. These findings further reveal the controversy in the conventional presentation of water dynamics and flow topography in the brain.
The conventional view on the brain nanodimentional interstitial space as a diffusion barrier fails to explain the new experimental results on water mobility and pathways in the brain parenchyma. In view of the failures of the conventional theory, there is an urgent call for the education reform in the area [24,25]. The debate around brain water metabolism is not just of a purely academic concern: on its solution depend many important practical issues.
Further doubts into validity of the conventional theory was introduced by the groundbreaking achievements in nanofluidics bringing new interdisciplinary knowledge into the area [9,10,26]. These achievements in nanofluidics laid grounds for development of the nanofluidic mechanism of brain water metabolism [9,10,24,26]. On the practical side this mechanism suggests, in particular, instead of residing to traumatic surgery, often with uncertain outcome, to use instead a AQP4-targeted drug therapy of brain water metabolism disorders [26].
The brain parenchyma incorporates the microvasculature, the neurons, and the glial cells, – all awash with a thin layer of ECF. The extracellular space, containing ECF, occupies, in total, about 20% of the brain volume [5,6]. The nanoconfined ECF surrounds all neural cells in the central nervous system. It presents an immediate external medium for the neural cells and is an integral part of brain metabolism, participating in various physiological functions.
The interconnected slits and channels with characteristic dimensions of 20-60 nm form an intricate network of the ECS continuum [27]. On the basis of its nanodimentionality, the water moiety there is classified as nanofluidic domain where fluid behaviour is governed by the principles of nanofluidics [8-10]. Notwithstanding this, a tendency still persist to employ a bulk-water approach to describe water behaviour in the nanodimentional ECS [3,7].
The results presented in this paper reveal some interesting features in water metabolism of the brain cortex. Thus, in the poorly vascularized Layer 1 there dominates water filtration with the penetrating arterioles serving as the main contributors to this process (Figure 2). The filtrated water is absorbed into the draining venule and the microvasculature in the lower layers of the brain cortex. As a result there is formed a steady flux of interstitial fluid directed from the top to the inner layers of the cortex. This finds its explanation in the gradual hydrostatic pressure drop observed in the microvasculature at the lower levels (Table 2). Blood supply to the neurons in the lower layers of the brain cortex is comparatively poor [28]. In view of this, an additional mass-transfer of nutrients with the interstitial fluid to the deeper layers of the brain cortex is beneficial.
The integral water flux within the model is positive. The interstitial space occupies 3.8 × 10-5 cm3 of the model volume. With the net positive water flux 7.72 × 10-8 cm3/s, the time necessary to completely replace the fluid in the interstitial space is 8.2 min. At the mean blood flow rate 1 cm/s [18,19], the total water flux, filtrated into the interstitial space, amounts to 2.8% of the volumetric blood flow rate of the four penetrating arterioles.
The cuboid model demonstrates the way the massive volume of the cerebrospinal fluid is formed outside the choroid plexus. More supporting information to this effect could be found in the literature [12,25]. This is in clear opposition to the orthodox concept of the choroid-plexus mechanism of CSF formation that, by now, has become an untenable dogma.
Water exchange in the oscillatory sections of the micro vessels is of a special interest. Contrary to the pulsatile flow patterns in FS and AS, water exchange in OS, between the blood and the brain parenchyma, is oscillatory. These oscillations proceed with the heart-beat-rate frequency. This oscillating pattern is conducive to realization of a fast feedback informational exchange between the blood ant the brain. OS moves along the blood microvessels, accordingly with the changes in topography of local hydrostatic pressure values (Figure 6). Full physiological import of the events taking place in and around OS is still to be elucidated.
An important role in brain water metabolism belongs to aquaporin AQP4. This water-conducting channel ensures fast water exchange between the blood and the extracellular fluid. AQP4 functions as a kinetically limiting step in water transfer between these two pools of water: the bulk water of the blood and the nano confined water in ECS. AQP4 is viewed as a molecular target for the drugs used in therapy of the conditions resulting from the watermetabolism- related disorders. At present, there is available a nomenclature of the AQP4-targeted drugs that modify activity of this wart channel and thus are capable to control water permeability of BBB. These drugs can find their practical use for therapy of brain edemas.
It should be observed that some drugs are widely used in clinical practice for therapy of various cases with doctors being un aware of their modifying effect on AQP4 activity and BBB permeability. Indiscriminate use of such drugs might result in appearance of untoward consequences and should be avoided. Fortunately, our current knowledge makes it possible to optimize the drug therapy without compromising brain water metabolism. The conventional approach to the flow of ISF and its driving mechanisms fails to account for many aspects of brain water metabolism [24]. Contrary to this, the concept of the brain nanofluidic domain provides deeper understanding of the workings of BWM.
For the first time a detailed water-flux topography in the brain cortex has been outlined. A fast fluid exchange, observed within the oscillatory sections of the microvasculature, is important for the mechanism of neurovascular coupling [29]. It is noteworthy that this oscillatory exchange, invisible from the conventional approach, has come into view only on introduction of the concept of the brain nanofluidic domain. The presented research deals primarily with the phenomenon of enhanced mobility of nano confined water due to the slipflow effect and the nanofluidic mass-transfer system [30-40].
The flow of ECF in its nano dimensional confinement defies our intuitive thinking about the expected behaviour of water within nanoscale network. However, this presents only a tip of an iceberg. There are other properties of nanoconfined water that make it so different from bulk water. For example, dielectric constant of nanoconfined water is an order of magnitude higher than that of bulk-water. Nano confinement profoundly modifies the energetics and the mechanisms of chemical reactions, enzymatic and catalyst performance. It impacts the polymer conformation. Thus, DNA, while globular in bulk water, unfolds itself in nanoconfined water. In view of this many physicochemical differences between nanoconfined and bulk water, they are considered completely different media.
Brain water fills the free-communicating spaces with a wide range of their characteristic dimensions. The geometry of these spaces scale down from the larger bulk-water-containing ones (e.g., the brain ventricles) to the nanodimentional geometry of the extracellular (interstitial) space containing nanoconfned water. In between, one finds micro-confined-water spaces (>1000 nm) and the extended-nanoconfined-water spaces (100-1000 nm). Before the advent of nanofluidics, the researchers were unaware of the compartment-size-dependent physicochemical properties of water and the hidden challenges coming henceforth.
The new interdisciplinary knowledge closes the gaps in this area. There appears a need for a revision of the experimental results where these differences have not been not taken into account. The new knowledge underscores the necessity of using a different approach to the events taking place in the spaces of nanometre-scale geometries. The confinement-sizedefined specific water properties and constraints are critically important and should be taken into account while considering biochemical and biophysical processes taking place in the brain, action of drugs and various issues related to brain physiology and pathology. In short, we should be keenly aware what waters we are navigating.
Apart from a promise of theoretical gains and deeper understanding the brain functions, the concept of the brain nanofluidic domain opens new ways to solution of a number of practical issues. It encompasses the therapy of brain edema, targeted delivery of drugs, ways of preventing cancer cell migration, control of beta-amyloid deposit formation, etc. It makes visible otherwise hidden events, opens new perspectives and promising venues for solving practical issues of brain physiology and pathology.
State basic research funding program # 20164106/182.
None.
The Author has declared that no competing interests exist.
Journal of Brain Research received 2 citations as per Google Scholar report