Mini Review - (2022) Volume 8, Issue 5
Received: 01-Sep-2022, Manuscript No. LDT-22-81081;
Editor assigned: 03-Sep-2022, Pre QC No. P-81081;
Reviewed: 17-Sep-2022, QC No. Q-81081;
Revised: 23-Sep-2022, Manuscript No. R-81081;
Published:
01-Oct-2022
, DOI: 10.37421/2472-1018.2022.8.161
Citation: Zhang, Xiaochen. “Alveolar Macrophage Propagation is controlled by Several Neutrophil Death Alleyway.” J Lung Dis Treat 8(2022):161.
Copyright: © 2022 Zhang X. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
The proliferation and self-renewal of alveolar macrophages (AM) are significant components of the microenvironment of lung tissue. The effect of immune cells, particularly neutrophils, on the homeostasis or functionality of AM is not fully understood. In this investigation, we used CXCL1 to promote neutrophil in vivo migration into bronchoalveolar lavage (BAL) fluid and lung, and we co-cultured these cells with AMs in vitro. It was discovered that BAL neutrophils (BAL-neutrophils) as opposed to BM-neutrophils limit AM growth. An analysis of data that was made accessible to the public revealed that there was significant molecular heterogeneity between blood and BM neutrophils and lung neutrophils. Unexpectedly, BAL-neutrophils from mice infected with the influenza virus PR8 (PR8-neutrophils) did not prevent the growth of AMs. Additionally, bulk RNA sequencing demonstrated that co-culturing AMs with PR8 neutrophils caused IFN- to be produced. In addition, BAL neutrophils from mice infected with PR8 altered the polarisation and phagocytosis of AMs and AMs co-cultured with BAL neutrophils had increased expression of metabolism- and ROS-associated genes. Combining inhibitors of various neutrophil death mechanisms reversed the inhibition of AM growth by BAL-neutrophils. Our findings imply that different neutrophil cell death processes control the proliferation of AMs. An alternative therapeutic approach for enhancing AM homeostasis in respiratory diseases may involve targeting neutrophil death.
Alveolar macrophages • Neutrophils • Influenza virus • Proliferation • Cell death • Inhibitors
Alveolar macrophages (AMs) are malleable cells that dwell in the alveolar milieu of lung tissue. As the first point of contact with the tissue microenvironment, AMs are distinct from other resident macrophages. In the pulmonary airspaces, AMs serve as sentinels and are crucial for the preservation of homeostasis, tissue healing, and immunological surveillance in the respiratory system. In the steady state, AMs are necessary for cellular debris clearance and the release of soluble mediators, both of which prevent host damage and safeguard pulmonary function. Pulmonary alveolar proteinosis (PAP), a syndrome brought on by an abnormal buildup of alveolar surfactant in the lung, can develop when AMs are absent or dysfunctional; in addition, ablation of AMs was also shown to increase in lung damage brought on by the flu virus. Therefore, maintaining AMs is essential for maintaining lung homeostasis. In terms of immunological surveillance, it has been demonstrated that AMs inhibit T cell growth, causing them to halt in the G0/G1 phase of the cell cycle.
Pulmonary dendritic cell (DC) maturation and T cell activation have both been found to be inhibited by AMs. By presenting transforming growth factor (TGF)-beta (RGA) and retinoic acid to CD4 T cells, AMs were reported to induce the development of Foxp3+ regulatory T cells (Foxp3+ Tregs), which reduced allergic airway inflammation. However, it has been demonstrated that the interferon (IFN) that pulmonary T cells secrete when infected with the influenza virus inhibits AM-mediated antibacterial defence. Neighboring alveolar epithelial cells' inflammatory responses may be controlled by AMs (AECs) Surfactant proteins A and D generated by AECs can also decrease AM immunological responses through cell-cell communication via cytokines. In a related study, it was discovered that the unique phenotype of lung basophils can influence AM maturation and function by combining lung IL-33 and GMCSF. However, it is unclear how other immune cell types, particularly those in the lung, affect AM self-renewal, development, and function [1].
Recruitment of neutrophils in bronchoalveolar lumen
5 g of recombinant murine CXCL1 was administered intranasally to anaesthetized C57BL/6 mice (rCXCL1, BioLegend, San Diego, CA, USA). A trachea incision was used to flush the airway five times with 1 mL cold, sterile BAL washes ([PBS, Hyclone] with 2% foetal bovine serum [FBS, Gibco] and 2 mM ethylenediaminetetraacetic acid [EDTA, Santa Cruz Biotechnology, Dallas, TX, USA]); the collected fluid was then centrifuged at 4°C, 800 g for five minutes. B After centrifuging for 5 minutes, the cell pellet was resuspended in ACK lysis buffer (0.15 M NH4Cl, 1 mM KHCO3, 0.1 mM Na2EDTA, pH 7.2) for 1 minute before the cells were suspended in MACS buffer. Using mouse anti- Ly6G, neutrophils were sorted to purify the neutrophils. UltraPure MicroBeads (Miltenyi Biotec, Teterow, Germany). BAL-neutrophils that express Ly6G were gathered. 0.4% Trypan Blue was used to measure cell number (Hyclone, Logan, UT, USA). Neutrophils from BAL from PR8-infected mice were obtained using anti-Ly6G MicroBeads after the mice were sacrificed three days after the infection [2].
RNA sequencing and analysis
We gathered AMs from AMs alone, AMs in co-culture with BAL-neutrophils, and AMs in co-culture with PR8-neutrophils. AMs' total RNA was extracted according to the manufacturer's instructions using a Trizol kit from Thermo Fisher Scientific. For bulk RNA-seq, two pools per genotype were employed. After quality assurance, the RNA sequencing library was created using highquality whole RNA (Agilent Bioanalyzer RIN of >7.0, Agilent Technologies, Santa Clara, CA, USA). The TruSeq RNA Sample Prep Kit v2 instructions for cDNA synthesis, end-repair, A-base addition, and ligation of the Illumina indexed adapters were followed (Illumina, San Diego, CA, USA). An Agilent Bioanalyzer DNA 1000 chip (Santa Clara, CA, USA) and Qubit fluorometry were used to measure the concentration and size distribution of the finished libraries (Invitrogen, Carlsbad, CA, USA) [3].
The sequencing of paired-end libraries on Base-calling was carried out with the DNBSEQ resequencing and PE 150 Kit (DNBSEQ A0, Beijing Genomics Institution, Shenzhen, China). In order to align paired-end RNA-seq reads, the mouse reference genome (GRCm38/mm10) was used. Using the NCBI mouse RefSeq gene model, pre- and post-alignment quality controls, gene level raw read count, and normalised read count (i.e., FPKM) were carried out using the RSeQC programme (v2.3.6) from Beijing Genomics Institution, Shenzhen, China. A weighted enrichment statistic and a log2 ratio metric for ranking genes were used in the Gene set enrichment analysis (GSEA) for functional analysis to find enriched gene sets utilising the hallmark collection of the Molecular Signatures Database (MSigDB), which contains upand downregulated genes. Information was provided to the GEO repository [4].
In both health and disease, neutrophils are essential for controlling innate and adaptive immune responses. Neutrophils often have a half-life of fewer than 24 hours. When neutrophils die, their cytotoxic components might be released, endangering the host and resulting in inflammatory or autoimmune illness. For instance, neutropenia brought on by bacterial or fungal infections has been treated with granulocyte transfusion and G-CSF. However, this therapeutic strategy is constrained by neutrophils' quick demise. Following infection with the influenza virus, there is a global neutrophil cell depletion that intensifies the inflammatory response and worsens the disease However, a slight reduction in neutrophils can promote an antiviral response. AMs and neutrophils were co-cultured in our study for more than In all tests, neutrophil mortality was noted after 24 hours (data not shown). We selected a few inhibitors to treat the co-culture of AMs and BAL-neutrophils in accordance with Fan et alfindings .'s. For instance, Q-VD-OPh, a pan-caspase inhibitor, inhibits caspase 3, 7, 8, 9, 10 and 13 to prevent apoptosis. NAC can improve neutrophil functions during acute pancreatitis and reduce neutrophil death in resting cells. DFO is a slow iron chelator that can stop NETs and ROS in neutrophils [5].
exhibit distinct traits. Our findings demonstrate that, particularly in light of the diverse ways of death, neutrophil life and death play a significant role in the proliferation and homeostasis of tissue-resident macrophages in pulmonary airspaces. Utilizing several inhibitors of neutrophil mortality in combination may restore AM proliferation and control AM homeostasis.
None
None.
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
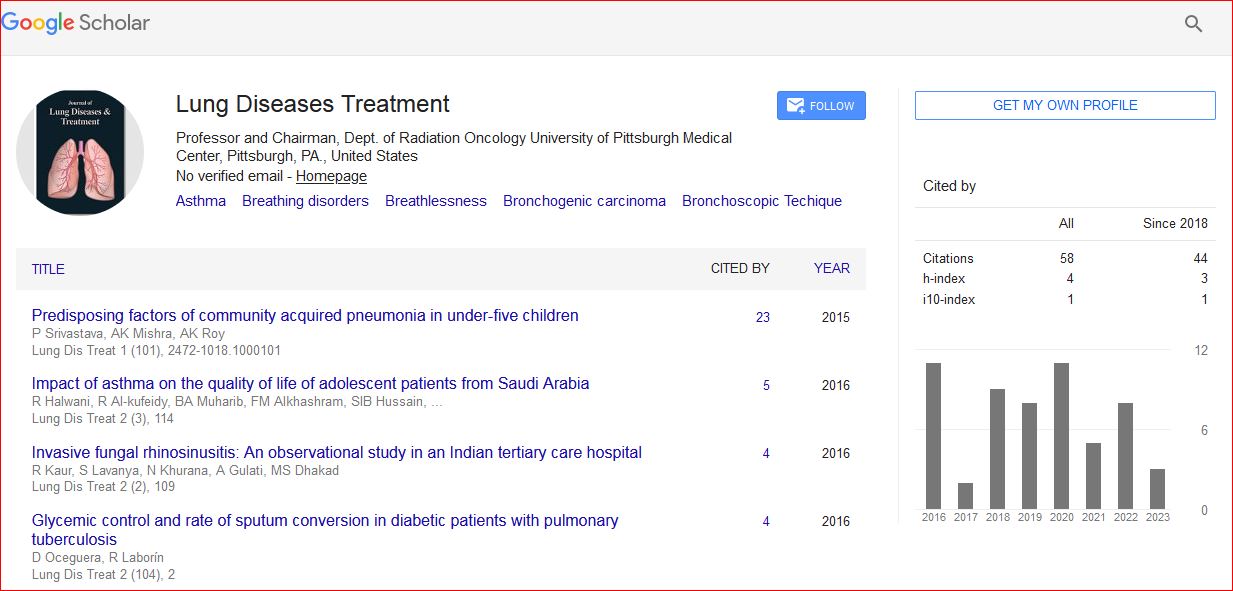
Journal of Lung Diseases & Treatment received 247 citations as per Google Scholar report