Original Research Article - (2022) Volume 0, Issue 0
Received: 15-Jun-2022, Manuscript No. VCRH-22-66274;
Editor assigned: 17-Jun-2022, Pre QC No. VCRH-22-66274, Pre Q C ;
Reviewed: 29-Jun-2022, QC No. VCRH-22-66274;
Revised: 08-Jul-2022, Manuscript No. VCRH-22-66274;
Published:
15-Jul-2022
, DOI: 10.37421/2736-657X.06.2022.004
Citation: Zhang, Hui, Haohui Huang, Rong Li and Lu Zhang "Broadly Potent Neutralizing Bi-specific Antibody against SARS-CoV-2." Virol Curr Res S1 (2022): 004
Copyright: © 2022 Zhang H, et al. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
The Omicron variant of Severe Acute Respiratory Syndrome Corona Virus-2 (SARS-CoV-2) is threatening our global efforts fighting the COVID-19 pandemic. As the vast majority of current countermeasures against SARS-CoV-2 are completely or markedly losing their effectiveness, including most of the clinically approved neutralizing antibodies, better and more efficacious novel agents are urgently needed. We have developed a bispecific antibody, BAT2022, which bonds simultaneously with high affinity to two non-overlapping epitopes on the Receptor-Binding Domain (RBD), competitively blocks the binding of RBD to human Angiotensin-Converting Enzyme2 (ACE2) and potently neutralizes SARS-CoV-2 and all of the variants tested. The IC50 values in the pseudovirus assay for Omicron (BA.1) and Delta are 30 pM and 50 pM, respectively. A mouse model of SARS-CoV-2, BAT2022 showed strong prophylactic and therapeutic effects. Prophylactically, a single administration of BAT2022 completely protected mice from bodyweight loss, as compared with up to 20% loss of body weight in placebo- treated mice, reduced the lung viral titers to undetectable in all mice treated with BAT2022 either prophylactically or therapeutically, as compared with around 1 × 105 pfu/g lung tissue in placebo-treated mice. Overall, Bispecific Antibody BAT2022 showed potent binding and neutralizing activity across a variety of SARS- CoV-2 variants and could be an attractive weapon to combat the ongoing waves of the Omicron pandemic.
COVID-19 pandemic • SARS-CoV-2 neutralization • Bio-Thera •Receptor-binding domain
The COVID-19 pandemic is continuously spreading around the world. As of December 2021, there were over 270 million confirmed cases and 5 million deaths across nearly 200 countries (https://COVID19.who.int/). The SARS- CoV-2 virus has continued to evolve throughout the pandemic [1-3]. Variants continue to emerge and some might become more contagious, virulent, or resistant to the current effective vaccines and neutralizing antibodies. Such variants are often referred to as Variants of Concern (VOC) [4-6]. Omicron is one such variant that is highly transmissible and resistant to most of our current countermeasures, such as vaccines and neutralizing antibodies [6-8].
Several neutralizing antibodies have demonstrated clinical efficacy and were approved by the Food and Drug Administration (FDA) under an Emergency Use Authorization (EUA) for the treatment of patients with mild to moderate COVID-19, especially for those with pre-existing medical conditions and are at higher risk of developing severe symptoms following infection [9]. Numerous neutralizing antibodies are currently under advanced clinical development phases and have shown promising efficacy. As the COVID-19 pandemic continues to spread, neutralizing antibodies, as a readily available therapeutic option, plays an important role in the fight against COVID-19 and hence, effectively protecting vulnerable people from getting infected or developing severe diseases following SARS-CoV-2 infection [10,11].
Most of the neutralizing antibodies target the RBD, which is important for cellular attachment and entry into host cells [12,13]. Unfortunately, mutations are inevitable for the SARS-CoV-2 coronavirus, just like many other viruses. Variants have emerged carrying mutations in the RBD that could potentially weaken the effectiveness of existing neutralizing antibodies or vaccines [14 -16]. Studies have consistently shown that some mutations in the RBD, such as K417N/T, L452R, E484K/Q, and N501Y can substantially reduce the strength of some neutralizing antibodies and vaccines [17, 18]. For instance, FDA has revoked the EUA for bamlanivimab monotherapy for COVID-19 due to the loss of efficacy against some of the currently circulating VOCs. Diamond and colleagues have recently shown that REGN10933, LY-CoV555, and 2B04 exhibited a marked or complete loss of neutralizing activity against variants like B.1.351, B.1.1.28, and viruses containing the E484K mutation [19]. Furthermore, studies have shown that monotherapy with a single antibody could lead to virus escape, both in vitro and in vivo [20-22]. Indeed, the current resurgence at an alarming speed of the COVID-19 pandemic in regions with very high vaccination rate has generated major concern around the world with numerous cases of breakthrough infections [23-25]. Therefore, novel antibodies that maintain their neutralization strength and breadth against the abovementioned resistant variants and possibly future emerging variants are urgently needed, which could offer extremely valuable and readily available countermeasures to combat the current wave of the COVID-19 pandemic caused mainly by VOCs.
In this study, we developed BAT2022, a fully-human bispecific antibody that binds two distinct epitopes on the RBD simultaneously, blocks the interaction between the RBD and ACE2, and potently neutralizes all currently (as of January 30, BAT2022) known VOCs of SARS-CoV-2, including Omicron, a known variant resistant to most of the clinical approved neutralizing antibodies, such as cocktails of bamlanivimab plus etesevimab, and casirivimab plus imdevimab, the two most widely used neutralizing antibodies cocktails. Therefore, BAT2022 could be a very attractive and potent therapeutic option to combat the current COVID-19 pandemic caused by Omicron.
Study design
This study was aimed to generate bispecific SARS-CoV-2 neutralizing antibodies resistant to mutations. To do so, we panned our proprietary fully- synthetic Intelligent Designed and Engineered Antibody Libraries (IDEAL), including one VH/VL-display phage library and one VHH-display phage library. The best candidate was selected based on high-affinity binding, potent neutralizing activity, and distinct binding epitopes. One candidate from each library was chosen to generate bispecific antibody BAT2022. BAT2022 was first comprehensively characterized in vitro for binding to mutant RBD or spike with mutations carried by VOCs and Variants of Interest (VOI), or mutations of residues key to interaction between ACE2 and RBD. Pseudovirus and live- virus neutralization assays were employed to assess the neutralizing activity of bispecific antibody BAT2022 against various variants. The in vivo efficacy of BAT2022 in both prophylactic and treatment settings was investigated in a well- established mouse model of SARS-CoV-2 [26]. The virus doses for challenge and timing of sample collection were selected based on a previous study. In a live virus-neutralizing assay, researchers were blinded to the identity of samples.
Cell lines
HEK293 (ACS-4500TM, ATCC) and African Green monkey kidney-derived VeroE6 cells (CRL-1587, ATCC) were cultured and passaged in Dulbecco's Modified Eagle Medium (DMEM) with 10% Fetal Bovine Serum (FBS). HEK293- hACE2 was constructed and sorted by Bio-Thera Solutions, Ltd.
Recombinant proteins
Biotinylated 2019-nCoV RBD, His, Avitag™ (SPD-C82E9, Acrobiosystems); SARS-CoV-2 RBD, His Tag (SPD-S52H6, Acrobiosystems); SARS-CoV-2 S1 protein (Ser 14-Arg 667), His Tag (S1N-S52H5, Acrobiosystems); SARS-CoV-2 S protein (R683A, R685A), His Tag, active trimer (SPN-C52H8, Acrobiosystems); SARS-CoV-2 RBD (N354D, D364Y), His Tag (SPD-S52H3, Acrobiosystems); SARS-CoV-2 RBD (W436R), His Tag (SPD-S52H7, Acrobiosystems); SARS- CoV-2 RBD (V367F), His Tag (SPD-S52H4, Acrobiosystems); SARS-CoV-2 RBD (L452R, E484Q) Protein (His Tag) ( 40592-V08H88ï¼?Sino Biological).
Surrogate assays for SARS-CoV-2-pseudovirus strains
Wildtype pseudovirus (GM-0220PV07, Genomeditech, Wuhan strain); Variant (D614G) (GM-0220PV14, Genomeditech); Variant (E484K)(GM- 0220PV35, Genomeditech); Variant (D614G, D936Y) (GM-0220PV19, Genomeditech); Variant (D839Y) (GM-0220PV6, Genomeditech); Variant (V483A) (GM-0220PV17, Genomeditech); Variant (D614G, A831V) (GM- 0220PV24, Genomeditech); Variant (W436R) (GM-0220PV26, Genomeditech); Variant (B.1.1.7/VUI-202012/01 del 145Y) (GM-0220PV33, Genomeditech); Variant (B.1.351/501Y.V2) (GM-0220PV32, Genomeditech).
Mice
Specific pathogen-free 6-10 weeks old female BALB/c mice were purchased from Hunan SJA Laboratory Animal Co. (Hunan, China) and bred in Animal Care Facilities of Guangzhou Customs Technology Center and Guangzhou Medical University. Mice were mildly anesthetized with brief exposure to isoflurane inhalant when needed.
SARS-CoV-2 viruses
The Accession number of wild-type strain used in the research is a clinical isolated SARS-CoV-2 virus 2019-nCoV/IQTC01/human/2020/Guangzhou (GenBank: MT123290.1). The SARS-CoV-2 beta variant (SARS-CoV-2/human/ CHN/20SF18530/2020 strain was isolated from an infected male individual and passaged on Vero E6 cells; the Accession number on National Pathogen Resource Center is NPRC2.062100001). The SARS-CoV-2 delta variant was offered by the Guangdong Provincial Center for Disease Control and Prevention.
Panning of phage display library
Our proprietary fully-synthetic IDEAL libraries contain one VH/VL-display phage library and one VHH-display phage library. The two libraries were panned separately on biotinylated recombinant SARS-CoV-2 RBD (SPD- C82E9, Acrobiosystems) in solution, In panning round 1, phages (1014 cfu) from each library were blocked with 3% Bovine Serum Albumin (BSA) in PBS/0.1% Tween-20 for 2 hours at room temperature, subsequently biotin-RBD was added and incubated for 1 h at room temperature. The phages were then captured by incubation with 50 μl Dynabeads MyOne Streptavidin (Invitrogen) for 30 min. Non-specific phages were eliminated by ten washes with PBS/0.1% Tween-20, followed by five washes with PBS for 5 min each. Bound phages were eluted with 10 mg/ml trypsin in PBS for 30 min, and trypsin was then inactivated with 10% fetal bovine serum. The eluted phages were re-infected into log-phase TG1 cells and amplified. A total of 4 consecutive panning rounds were performed with decreasing amounts of antigen for each subsequent round, 10 ug, 2.5 ug, 0.5 ug, and 0.1 ug of antigen was used for round 1 to 4, respectively. After 4 rounds of panning, VH/VL and VHH DNA sequences were pool cloned from the phage displaying vector into the prokaryotic expression vector, then transfected into BL21 Escherichia coli. Single colonies were picked and expressed, soluble scFv or VHH in the supernatant was assessed by ELISA for RBD-binding.
Enzyme-Linked Immunosorbent Assay (ELISA)
One ug/ml His-tagged Spike RBD protein of SARS-CoV-2, as well as mutated S1 domains, RBDs, or ectodomain of trimeric Spike were immobilized onto 96-well plates (9018, Corning) overnight at 4°C, plates were blocked with 3% BSA in Phosphate Buffered Saline with Tween 20 (PBST) for 2 hours at 37°C. Samples were serially 3-fold diluted and added 100 ul per well into blocked plates, incubated for 1 hour at 37°C. Bound antibodies were detected with peroxidase-conjugated goat anti-human kappa light chains antibody (A7164- 1ML, Sigma). 100 ul TMB (Tetra Methylbenzidine, Biopanda TMB-S-001) was added per well to develop. Experiments were conducted in duplicates. EC50 values were determined by 4-parameter logistic regression analysis using Softmax Pro 7.
ACE2 Receptor-binding blocking ELISA
One ug/ml His-tagged spike RBD protein of SARS-CoV-2 (SPD-C52H3, Acrobiosystems) was immobilized onto 96-well plates (9018, Corning) overnight at 4°C, plates were blocked with 3%-5% BSA in PBST for 2 hours at 37°C. Serially 3-fold diluted BAT2022 was added into the blocked plate, incubated for 1 hour at 37°C. Biotinylated human ACE2 protein (AC2-H82E6, Acrobiosystems) was added to the plate with antibody diluted inocula to the final concentration of 25 ng/ml, further incubated for 1 hour at 37°C. The remaining biotinylated human ACE2 binding to the RBD coating on the plate was detected using HRP-labelled streptavidin (016-030-084, Jackson Immunoresearch). The plate was developed with TMB (Tetra Methylbenzidine, Biopanda TMB-S-001). Absorbance at 450 nm was measured on SpectraMax Plus Absorbance Microplate Reader (Molecular Device, CA). IC50 values were determined by 4-parameter logistic regression analysis using Softmax Pro 7.
Antibody and recombinant RBD expression and purification
The BAT2022 coding sequences of heavy and light chains were cloned to expression plasmids, respectively. HEK293 cells were transiently co-transfected at a ratio of 1:2 (H:L) with polyethylenimine (PEI; 49553-93-7, Polyscience) according to the manufacturer’s instruction. The supernatants were harvested at Day 7 post-transfection and purified by protein-A affinity column. For the expression of recombinant alanine mutants, we designed and synthesized 10 RBD DNA sequences (GENERAL BIOL), each one containing one specific alanine mutant of 10 critical residues related to interaction with ACE2 and cloned into an expression vector with an Fc tag. Transiently transfected to HEK293 cells to obtain recombinant RBD alanine mutants and purified by protein A columns.
Affinity measurement of antibodies by Surface Plasmon Resonance (SPR)
The SPR experiments were all conducted with a Biacore T200 system (GE Healthcare); All assays were performed with a Sensor Chip Protein A (GE Healthcare), with an HBS EP+ running buffer (0.1M HEPES, 1.5M NaCl, 0.03M EDTA supplemented with 0.005% vol/vol surfactant P20 at 25â??.) To determine the affinities of nanobody VHH18, human Immunoglobulin G1 (IgG1) antibody 2F8, and BAT2022 to SARS-CoV-2 RBD-His tag, spike trimer-His tag and other S1-His tag variants, antibodies were immobilized onto the sample flow paths of the sensor Protein A chip. The reference flow cell was left blank. SARS-CoV-2 RBD-His tag, or spike trimer-His tag or other S1-His tag variants was injected over the above-mentioned flow paths at a range of five concentrations prepared by 2-fold serial dilutions started at 50nM, at a flow rate of 30 ul/min, with an association time of 75s and a dissociation time of 180s. HBS EP+ running buffer was also injected using the same program for background subtraction. All the data were fitted to a 1:1 binding model using Biacore T200 Evaluation Software 3.1.
Epitope binning by in-tandem forte bio assay
Epitope analysis for 2F8 and VHH18 was carried out by in-tandem Bio Layer Interferometry (BLI) via Octet QKe (ForteBio) according to the manufacturer’s instruction. His, Avitag SARS-CoV-2 RBD protein (SPD-C82E9, Acrobiosystems) was diluted to a final concentration of 400 nM in kinetics buffer (1 × PBS, 0.01% Tweens-20) and immobilized onto Strepavidin biosensor. The sensor was saturated with the first antibody, either 2F8 Fab or VHH18-His. Subsequently, the above bioprobes were flown over with the different second antibodies, either VHH-his or 2F8 Fab, respectively.
Pseudo virus neutralization assay
HEK293 was transfected with human ACE2 cDNA cloning vector (HG10108-M, Sino Biological), and sorted with BD FACJazz cell sorter to get monoclonal cell line, HEK293-hACE2. BAT2022 was serially 3-fold diluted in duplicate and incubated with SARS-CoV-2 pseudovirus or other variant pseudoviruses which were diluted according to manufacturer’s instruction for 1 hour at 37°C. HEK293-hACE2 was detached and then added to the pseudo virus-antibody mixtures. No antibody was used as positive control, and no pseudo virus was used as negative control. After 48 hours of incubation at 37°C in 5% CO2 , neutralization potencies were quantified in luciferase assay measured with Bio-Lite Luciferase Assay solutions (DD1201-03, Vazyme). The values were read on microplate reader SpectraMax M3 (Molecular Devices). The Half-Maximal Inhibitory Concentration (IC50) and 90% of Maximal Inhibitory Concentration (IC90) were determined by four parameter logistic regression.
Focus Reduction Neutralization Test (FRNT)
Bispecific antibody BAT2022 was serially 3-fold diluted in DMEM and incubated with an equal volume of SARS-CoV-2 wildtype (Wuhan strain) or delta variant containing 200 PFU for 1 hour, at 37°C. The mixtures were then added to Vero E6 monolayers in a 96-well plate in triplicate and incubated for 1 hour at 37°C in 5% CO2. The inocula were removed and added 100 ul pre-warmed 1.6% (w/v) CMC (Carboxy Methyl Cellulose) in MEM containing 2% FBS per well, further incubate for 24 hours at 37°C in 5% CO . Cells were then fixed with 4% Para Formaldehyde (PFA) and permeabilized using 0.2% Triton X-100. Cells were tested with a rabbit anti-SARS-CoV-2 nucleocapsid protein polyclonal antibody (Cat. No.: 40143-T62, SinoBiological, Inc.), and an HRP-labelled goat anti-rabbit as a secondary antibody (111-035-003, Jackson ImmunoResearch). The foci were developed by Trueblue peroxidase substrate and results were read on Immuno-ELISPOT (CTL ImmunoSpot UV). IC50 values were determined using four parameter logistic regression (GraphPad Prism). Experiments were conducted under the standard operating procedures of the approved Biosafety Level-3 laboratory.
Animal experiment
All animal experiments were performed under the relevant ethical regulations regarding animal research. The mice experiments for in vivo efficacy were conducted in the Animal Care Facilities of Guangzhou Customs District Technology Center and Guangzhou Medical University. All protocols were approved by the Institutional animal care and use committees of guangzhou customs district technology center and guangzhou medical university. (approval numbers: 202092 ). All the live virus experiments were conducted in the Biosafety Level-3 laboratory of Guangzhou Customs District Technology center following standard operating procedures. This study is reported per the ARRIVE guidelines.
BALB/c mice were mildly anesthetized with isoflurane and intranasally transduced with 2.5 × 109 PFU of Ad5-ACE2 in 75ul DMEM. Four days after Ad5-ACE2 transduction, mice were infected via i.n. route with 1 × 105 PFU SARS-CoV-2 virus. For prophylaxis, mice were dosed with 1 mg antibody either via i.p or i.n. route, 24 hours before virus infection and for treatment, mice were dosed with 1 mg antibody via i.p. route, 18 hours post virus challenge, PBS was used as placebo. Animals were monitored daily and body weight was recorded daily. Lung tissues were harvested at 3 DPI for viral titers measurement by Focus Reduction Neutralization Test (FRNT). About 140 mg of lung tissue from each mouse (n=3, or 4) was homogenized to 1 ml PBS, 50 ul lung homogenate supernatants were used for viral titers.
Statistical analysis
The neutralization potency of BAT2022 was defined as percent inhibition, and the four parameter logistic regression model fit was used to calculate the IC50. The Half-maximum Effective Concentration (EC50) was calculated using 4-parameter logistic model.
Characterization of monoclonal neutralizing antibody 2F8 and VHH18
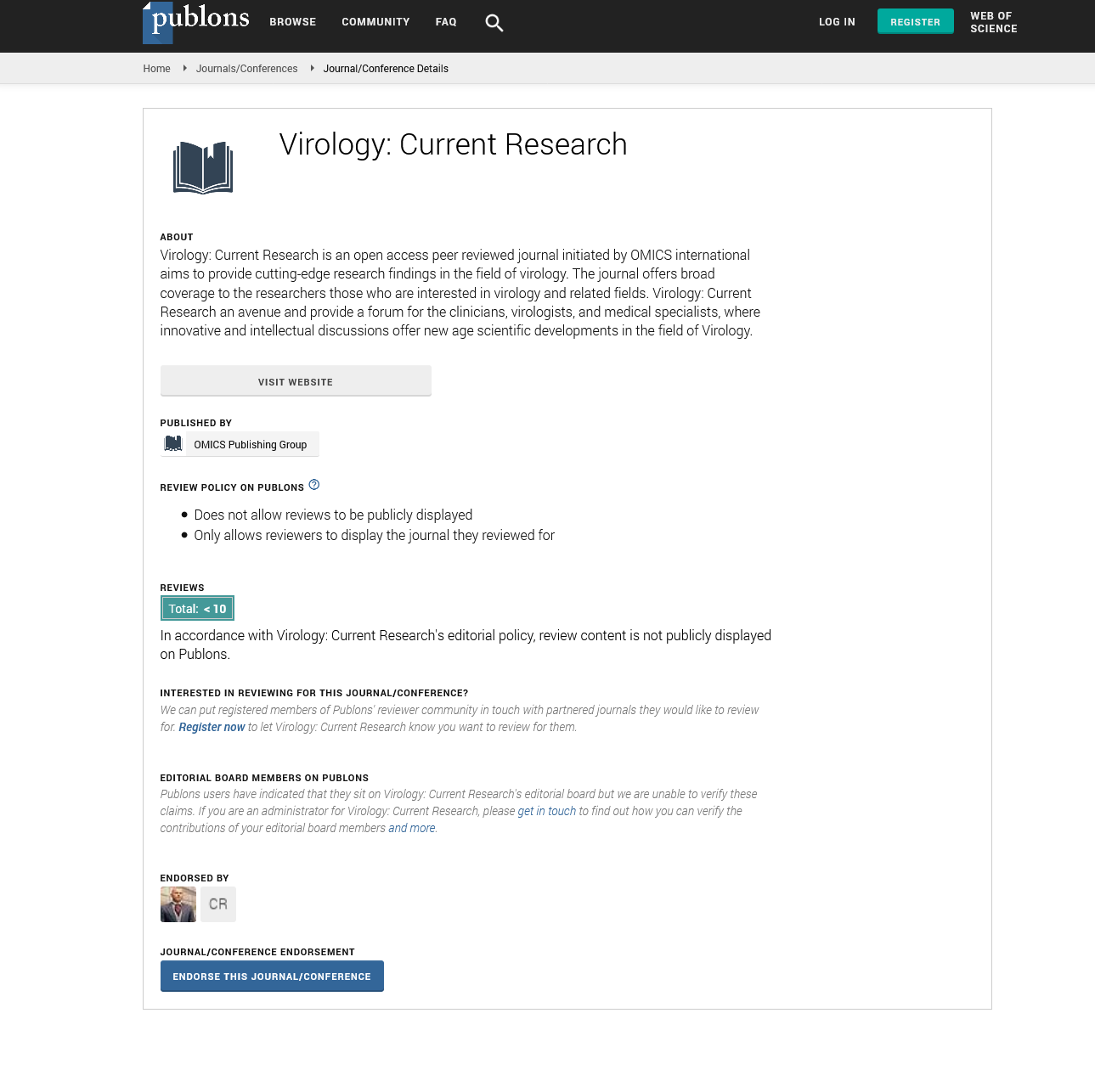
Spike RBD is the major target for SARS-CoV-2 neutralizing antibodies. In order to isolate neutralizing antibodies against SARS-CoV-2, we used recombinant RBD of SARS-CoV-2 spike protein as antigen, panning from Bio- Thera’s proprietary IDEAL synthetic VH/VL and nanobody fully human phage library separately. After four rounds of panning, Soluble Single-chain Fragment Variable (ScFv) or nanobody candidates were first assessed for RBD binding in Enzyme-Linked Immunosorbent Assay (ELISA). Positive clones were then converted to full IgG or VHH-Fc and assessed for their abilities to block RBD binding to ACE2 and to neutralize SARS-CoV-2 pseudovirus, and for epitope binning. The two best lead antibodies i.e., 2F8 and VHH18, isolated from the VH/VL library and nanobody library, respectively were chosen to generate the Bispecific Antibody, BAT2022. Both 2F8 and VHH18 bind specifically to Recombinant RBD and the ectodomain of the trimeric spike of SARS-CoV-2 with high affinity in ELISA, with EC50 values of 130 pM and 150 pM for RBD (Figure 1a), 130 pM and 190 pM for spike trimer (Figure 1b), respectively. In addition, 2F8 can completely block the binding of RBD to recombinant human ACE2, with an IC50 value of 190 pM compared to VHH18, which is known to partially inhibit the interaction between RBD and recombinant human ACE2 (Figure 1c). Moreover, both 2F8 and VHH18 can potently neutralize SARS- CoV-2 pseudovirus with IC50 values of 45 pM and 1600 pM, respectively (Figure 1d). Significantly, 2F8 and VHH18 can bind distinct non-overlapping epitopes on RBD as determined by Fortebio in-tandem assay. When immobilized RBD was first loaded with saturated 2F8, VHH18 was still able to bind RBD completely, indicating that 2F8 and VHH18 can bind RBD simultaneously and 2F8 and VHH18 bind distinct epitopes on RBD (Figures 1a-1e).

Figure 1a. Characterization of 2F8 and VHH18in vitro. Binding of 2F8 and VHH18 to recombinant RBD in ELISA. Data are mean ± S.D. of technical duplicates. EC50 values are shown on the graph. Note: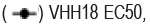
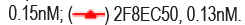

Figure 1b. Characterization of 2F8 and VHH18 in vitro. Binding of 2F8 and VHH18 to recombinant spike ectodomain trimer in ELISA. Data are mean ± S.D. of technical duplicates. EC50 values are shown on the graph. Note:
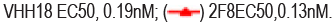

Figure 1c. Characterization of 2F8 and VHH18 in vitro. Blocking of RBD binding to recombinant human-ACE2 in ELISA. 

Figure 1d. Characterization of 2F8 and VHH18 in vitro. Neutralization activity against SARS-CoV-2 pseudovirus. Note:

Figure 1e. Characterization of 2F8 and VHH18in vitro. Epitope binning of 2F8 and VHH18 by in-tandem ForteBio. RBD was immobilized on chip followed by saturated 2F8, subsequently, VHH18 or 2F8 was loaded. VHH18 binds RBD in the presence of saturated 2F8, 2F8 binds RBD in the presence of saturated VHH18, indicating that VHH18 and 2F8 have distinct epitopes on RBD. 
Binding characterization of bi-specific neutralizing antibody BAT2022
Cocktails of neutralizing antibodies against SARS-CoV-2 have been successfully applied to combat viral escape in clinical settings. A bi-specific antibody can combine the advantages of cocktails into a single molecule. In our study, we chose 2F8 and VHH18 to generate the bi-specific Neutralizing Antibody BAT2022. As described above, 2F8 and VHH18 have non-overlapping distinct epitopes on RBD and both show activity against SARS-CoV-2. In order to prepare bi-specific Antibody BAT2022, VHH18 was linked to the C-terminal of the Fc of 2F8 (Figure 2a). The Binding Activity of BAT2022 was first assessed by ELISA. During our study, we noted that BAT2022 was able to bind strongly to both RBD and spike-trimer of SARS-CoV-2, with EC50 values of 270 pM and 260 pM, respectively (Figure 2b). Most neutralizing antibodies block SARS-CoV-2 infection by inhibiting the binding of RBD to ACE2. The activity of BAT2022 to inhibit the binding of RBD to ACE2 was assessed by ELISA. As shown in Figure 2c, BAT2022 strongly inhibited the binding of ACE2 to RBD (IC50=180 pM) and was then evaluated for binding to occurring SARSCoV- 2 variants tested, including mutations such as K417N/T, E484K, N501Y, L452R, and D614G, carried by VOC such as alpha, beta, gamma, delta, and Omicron (BA.1), with IC50 values ranging from 200 to 280 pM, indicating that these mutations in the RBD did not significantly impact the binding of BAT2022 (Table 1); in addition, we have also assessed the binding of BAT2022 to alanine- scan mutants of ten critical residues of RBD, including D405A, K417A, K444A, Y449A, F456A, K458A, F486A, F490A, V503A, Y505A, related to interaction with ACE2 in ELISA. BAT2022 bound strongly to all alanine mutants (Table 2), despite the fact that 2F8, one of the parental antibodies of BAT2022, completely lost binding to variant V503A (data not shown). Furthermore, the F486 mutation has been shown to reduce the effectiveness of casirivimab (F486I, F486V), bamlanivimab (F486V), and etesevimab (F486V) [27]. The F490S is one of the two (F490S, L452Q) mutations carried in RBD by lambda variant, yet, neither F486A nor F490A has an impact on BAT2022 binding. Together, these results suggest that BAT2022 could be effective against SARS-CoV-2 for all VOCs currently known. Not only this, BAT2022 could potentially be effective for future variants too (Figures 2a-2c) (Tables 1 and 2).
| Antigen | Variant | EC50 (nM) |
|---|---|---|
| Wildtype (Wuhan strain) | NA | 0.26 |
| HV69-70 deletion, Y144 deletion, N501Y,A570D, D614G,P681H | Alpha | 0.25 |
| K417N, E484K, N501Y, D614G | Beta, Gamma | 0.22 |
| L452R, T478K | Delta | 0.26 |
| G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H | Omicron (BA.1) |
0.27 |
| W436R | 0.28 | |
| N354D,D364Y | 0.2 | |
| V367F | 0.26 | |
| N501Y | 0.25 | |
| K417N | 0.25 | |
| D614G | 0.22 |
Note: EC50 values are the means of two technical duplicates
| Mutation | EC50 (nM) |
|---|---|
| D405A | 0.22 |
| K417A | 0.26 |
| K444A | 0.24 |
| Y449A | 0.28 |
| F456A | 0.37 |
| K458A | 0.22 |
| F486A | 0.23 |
| F490A | 0.22 |
| V503A | 0.19 |
| Y505A | 0.21 |
| Wildtype (wuhan strain) | 0.2 |
Note: EC50 values are the means of two technical duplicates

Figure 2a. Characterization of BAT2022in vitro. Schematic structure of BAT2022.

Figure 2b. CCharacterization of BAT2022in vitro. Binding of BAT2022 to recombinant RBD or ectodomain spike trimer in ELISA. Data are mean ± S.D. of technical duplicates. EC50 values are shown on the graph. 
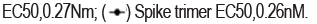

Figure 2c. Characterization of BAT2022in vitro. Binding of BAT2022 to recombinant RBD or ectodomain spike trimer in ELISA. Data are mean ± S.D. of technical duplicates. EC50 values are shown on the graph.

The binding kinetics of BAT2022 was measured by Surface Plasma Resonance (SPR). Bispecific Antibody BAT2022 was immobilized to Protein-A sensor and the affinities to RBD or spike ecto-trimer were measured. Consistent with the results of ELISA, BAT2022 bound with very high affinity to both the RBD and spike trimer of wild-type SARS-CoV-2, with Equilibrium Dissociation constant (KD) values of 45.9 pM and 47.7 pM, respectively. In addition, BAT2022 also bound mutant RBD or S1 with mutations found in all major VOCs reported so far with high affinities, KD values ranging from 1060 pM to 59 pM, which is 5 to 10-fold better than the benchmark neutralizing antibody 10933 for all variants tested (Table 3) [20]. Notably, BAT2022 has an affinity of 60.5 pM for the Omicron spike (Table 3).
| Strains | Antigen | Mutation | KD (M) |
|---|---|---|---|
| WT | RBD | NA | 4.59E-11 |
| WT | Spike Trimer | NA | 4.77E-11 |
| Alpha | Spike-S1 | HV69-70 deletion, Y144 deletion, N501Y, A570D, D614G, P681H | 4.02E-10 |
| Beta | Spike-S1 | L18F, D80A, D215G, LAL242-244 deletion, R246I, K417N, E484K, N501Y, D614G | 1.06E-09 |
| Gamma | Spike-S1 | L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, D614G, H655Y | 8.10E-10 |
| Delta | RBD | L452R, T478K, E484Q | 1.86E-11 |
| B.1.617.1 | RBD | L452R, E484Q | 5.97E-11 |
| Omicron (BA.1) |
RBD | G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H | 1.16E-10 |
| Omicron (BA.2.12.1) |
RBD | G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452Q, S477N, T478K, E484A, Q493R, Q498R, N501Y, Y505H | 1.15E-10 |
| Omicron (BA.1) |
Spike Trimer | A67V, Δ69-70, T95I, G142D, Δ143-145, Δ211, L212I, ins214EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F | 6.05E-11 |
Note: KD- equilibrium dissociation constant
BAT2022 potently neutralizes SARS-CoV-2 infection
The neutralizing activity of BAT2022 against SARS-CoV-2 was assessed using both pseudovirus and live virus assays. The neutralizing potency of BAT2022 was tested against luciferase reporter viruses, pseudo-typed with SARS-CoV-2 spike protein. The results of our study show that BAT2022 exhibited highly potent neutralizing activity against the wild-type and all mutant pseudovirus carrying mutations found in the past and the VOC designated in the past, such as alpha, beta, gamma, delta, and Omicron (Figure 2d and Table 4), with IC50 values ranging from 20 pM to 160 pM, IC90 values from 84 pM to 510 pM, including mutations known to be resistant to etesevimab (K417N) or bamlanivimab (E484K) [27]. Notably, BAT2022 showed exceptional potency for Omicron pseudovirus, with an IC50 value of 30 pM. In addition, the neutralizing activity of BAT2022 was tested against SARS-CoV-2 live virus by FRNT assay. It was observed that BAT2022 showed exceptionally potent neutralizing activity against wild-type live SARS-CoV-2, with an IC50 value of 26.4 pM and an IC90 value of 114 pM. Furthermore, BAT2022 also showed extremely potent neutralizing activity against the beta (IC50=13.3 pM) and delta variants (IC50=88.6 pM) as depicted in Figure 2e and Table 5(Figure 2c-2e) (Tables 4 and 5).
| Strains | IC50 (nM) | IC90 (nM) |
|---|---|---|
| Wild type | 0.13 | 0.325 |
| Alpha | 0.03 | 0.17 |
| Beta | 0.02 | 0.16 |
| Delta | 0.055 | 0.21 |
| Omicron (BA.1) | 0.03 | 0.12 |
| E484K+K417N+N501Y | 0.093 | 0.24 |
| E484K | 0.154 | 0.45 |
| K417N | 0.16 | 0.51 |
| D614G | 0.029 | 0.084 |
| N501Y, D614G | 0.028 | 0.19 |
| N354D, D364Y | 0.045 | 0.18 |
Note: IC50 = concentration inhibiting maximal activity by 50%; IC90 = concentration inhibiting maximal activity by 90%.
| Variant | IC50 (pM) | IC90 (pM) |
|---|---|---|
| Wild type | 26.4 | 114 |
| Beta | 13.3 | 35.4 |
| Delta | 88.6 | 259 |
Note: IC50 = concentration inhibiting maximal activity by 50%; IC90 = concentration inhibiting maximal activity by 90%.

Figure 2d. Characterization of BAT2022in vitro. Neutralization potency on SARS- CoV-2 pseudovirus. Data are mean ± S.D. of technical duplicates. 
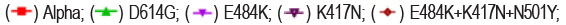
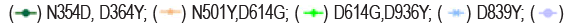
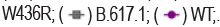

Figure 2e. Characterization of BAT2022in vitro. Neutralization potency on SARS-CoV-2 live virus. Data are mean ± S.D. of technical duplicates. IC50 values are shown on the graph. 
The in vivo efficacy of BAT2022 was assessed in a mouse model of SARS-CoV-2. In this model, mice were first transduced intranasally with an Adenovirus that expresses Human ACE2 (AdV-hACE2), which resulted in the expression of hACE2 in mice alveolar and airway epithelium, leading to the susceptibility of mice to be infected with SARS-CoV-2 live virus [26]. To assess the prophylactic efficacy of BAT2022, animals were treated with a single dose of 1 mg of BAT2022, or PBS one day before intranasally challenging with 1 × 105 Plaque-Forming Unit (PFU) of SARS-CoV-2 live virus (Figure 3a). As previously reported, substantial rapid weight loss of up to 20% at 4 Days Post-Infection (DPI) was observed in all 5 animals treated with PBS. By contrast, mice treated with BAT2022 prophylactically either via intranasal (i.n.) or intraperitoneal (i.p) route had no significant loss of body weight during the entire experiment (Figure 3b). The viral titers were determined in the whole lung tissue homogenates at 3 dpi by FRNT assay. The viral titers in the lungs were below the detection limit in all mice receiving BAT2022 prophylactically either via i.n or i.p route, compared with about 1 × 105 pfu/g lung tissue in all mice treated with PBS, indicating that BAT2022 could abolish virus replication in the lungs (Figure 3c). These results indicated that BAT2022 can block the establishment of virus infection when given prophylactically, suggesting that BAT2022 is efficacious even when administered as prophylaxis both i.n and i.p routes, as shown by the undetectable viral load in the lungs and no weight loss was reported 9 (Figures 3a-3c).

Figure 3a. In vivo efficacy of BAT2022 in mouse model of SARS-CoV-2. Schematic overview of study design. Animals were administered i.p. or i.n. a single dose (50 mg/kg) of BAT2022, or PBS. Animals were treated 24 hours before or 18 hours post virus challenge for prophylactic and therapeutic, respectively. Body weight was recorded daily for 11 days. 

Figure 3b. In vivo efficacy of BAT2022 in mouse model of SARS-CoV-2. Body weight changes of animals. Data are presented as mean ± SD. (n=5). Note: 

Figure 3c. In vivo efficacy of BAT2022 in mouse model of SARS-CoV-2. Whole lung virus titer at 3 dpi as assessed by FRNT assay. Each dot represents an individual animal (n=3, or 4). Dotted line indicates the detection limit of the FRNT assay. 
Therapeutic efficacy in a mouse model of SARS-CoV-2
The efficacy of BAT2022 was then assessed in the same mouse model in a therapeutic setting. Mice were treated i.p with a single dose (1 mg/mouse) of BAT2022, 18 hours after challenging with 1 × 105 plaque-forming units (PFU) of SARS-CoV-2 virus. Because viral replication and disease progression occurred very rapidly in mice challenged with 1 × 105 PFU of virus in this model, far exceeding the time course of COVID-19 in clinical. Nevertheless, a significant therapeutic benefit was observed in mice treated with BAT2022 [26]. The viral titers in the lung at 3 DPI were undetectable in all animals treated with BAT2022, as compared with ∼1 × 105 FFU/lung in all animals receiving PBS (Figure 3c), demonstrating that BAT2022 can constrain virus replication in vivo even when a virus infection was vigorously spreading. Even though weight loss was not prevented, due possibly to the rapid progress of the disease setting, animals receiving BAT2022 recovered faster than did PBS-treated mice, the difference is statistically significant at both day 10 (p=0.027) and day 11 (p=0.036). BAT2022-treated mice completely regained their body weight loss by 11 dpi whereas PBS-treated animals did not (Figure 3b). In summary, these in vivo results demonstrate that BAT2022 is efficacious against SARS-COV-2 infection in mice as a prophylactic or therapeutic treatment and that BAT2022 could be a promising candidate for the prevention and treatment of COVID-19 induced by SARS-COV-2 wild-type and possibly effective against other variants including Omicron BA.1 and BA.2.12.1 based on data from in vitro binding and pseudovirus neutralization assay.
In this study, a bispecific monoclonal antibody, BAT2022, was derived from two antibodies with distinct non-overlapping epitopes on RBD, which combine the advantages of two antibodies into a single molecule, retains potency and breadth. The evidence reported by the study shows that BAT2022 possess potent RBD-binding and virus-neutralizing activity across all of the reported VOC existed in the past, including alpha, beta, gamma, delta, and Omicron (BA.1) in particular, which is highly and broadly resistant to not only most of the clinically approved neutralizing antibodies but also most of the vaccines used around the globe.
It is noted that BAT2022 has demonstrated potent in vivo efficacy against SARS-CoV-2 in mice both prophylactically and therapeutically, protecting animals from disease progression and abolishing virus replication in the lung. When given prophylactically, animals were completely protected from bodyweight loss. Our results showed that BAT2022 can prevent infection in an animal model when given via i.n. or i.p. route. It would be interesting to evaluate how long the protection can last. Even though weight loss was not prevented in a therapeutic setting, animals receiving BAT2022 did regain weight loss faster than placebo-treated animals did. The previous study has shown prevention of weight loss in a similar mouse model, but with significantly less viral load (1 × 104 PFU vs. 1 × 105 PFU) and earlier dosing time point (12 hours vs. 18 hours) post virus challenge [21]. These results together with others’ findings, suggest that disease progression in this model is virus-load and treatment dosing-time post infection sensitive.
Our data suggest that a bispecific antibody might likely have broader spectrum and be more effective in inhibiting host-virus interactions and prevention of viral escape than single antibody alone. Studies have consistently shown that a single neutralizing monoclonal antibody might be more susceptible to viral mutant escape [28]. For instance, recently reported monoclonal antibody 2B11 showed potent neutralizing ability against wild-type SARS-CoV-2, yet, its neutralization activity against B.1.351 or P.1 pseudovirus was significantly impaired [29]. Therefore, cocktails of two or more antibodies are widely used to counteract virus escape. As a Bispecific Antibody, BAT2022 showed strong binding to variant even to which its parental antibody did not bind. Physically connecting the two non-over-lapping epitope-binding antibodies likely increases overall association constants (Kon values) and decreases overall dissociation constants (Koff values), thus increasing the affinity and also avidity of the bispecific antibody via its increased valency in a single molecule. A previous study has also demonstrated that biparatopic bispecific antibody has significantly higher affinity and stronger neutralizing activity against SARS-CoV-2 [30-33]. Overall, a single molecule with bispecificity and multi-valency will likely have better efficacy and a broader spectrum against mutations.
A major limitation of the current study is the lack of structural characterization of BAT2022 in complex with RBD and/or spike. Such information would be valuable to predict more precisely how BAT2022 acts, whether BAT2022 binds to the “up” or “down” or both conformers of the RBD on the S protein and whether the contact residues of BAT2022 on RBD have a high or low frequency of mutation. Epitopes with very low mutation frequency might indicate more resistance to virus escape. In addition, even though in vitro studies have shown that BAT2022 has potent and broad neutralizing activity against all variants tested, in vivo study has been done with wild-type SARS-CoV-2 only.
In conclusion, BAT2022 maintains strength and breadth across all of the reported VOC designated in the past (alpha, beta, gamma, delta, and Omicron (BA.1)) and a wide variety of variants with mutations known to compromise the effectiveness of neutralizing antibodies and vaccines currently used in the clinic. BAT2022 would be an attractive solution to the ongoing resurgence of the COVID-19 pandemic caused mainly by VOC, such as delta and Omicron, and possibly to potential future newly emerging variants and warrants evaluation in human clinical studies.
We are grateful to our colleagues at Bio-Thera Solutions for their support during the study, especially Chunping Deng, Jian Ma, Yanli Liu, Wengrong Huang, Chao Qin, Junjie Huang, Zhencheng Chen, Chuang Li. We thank Bert Thomas for reviewing this manuscript.
Statements and Declarations
Funding
This study was funded by Bio-thera Solutions.
Authors’ contribution
H.Z., H.H., Z.W., J.L., J.C., H.S., D.Z., Z.S., S.Z., C.L. constructed the IDEAL libraries and isolated the antibodies under the supervision of H.Z., J.Y., S.L., and X.H. R.L., L.Z., and Y.Y. performed the pseudovirus and live virus- neutralizing assay under the supervision of X.L. and H.Z. S.L. participated in the design of the nano-IDEAL library. S.P. and H.Y. expressed and purified recombinant alanine-scan mutants. L.W. and C.D. analyzed the biochemical and biophysical properties of the selected antibodies under the supervision of C.L. Y.W. and L.Z. performed the in vivo efficacy study in mice. H.Z., S.L., and X.H. conceived the project and wrote the manuscript.
Conflicts of interests
Hui Zhang, Haohui Huang, Zhiwei Wang, Shide Liang, Jiaping Li, Junyou Chen, Ziqi Su, Huafei Su, Li Wang, Chunping Deng, shujun Pei, Shenhua Zhu, Chan Li, Haitao Yue, Cuihua Liu, Jinchen Yu, Shengfeng Li, and Xianming Huang are employees and/or stockholders of Bio-Thera Solutions. This study was funded by Bio-Thera Solutions.
Data and materials availability
All processed data are available in the main text and all raw data are available upon request.
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Google Scholar] [Pubmed]
Virology: Current Research received 187 citations as per Google Scholar report