Image Article - (2021) Volume 11, Issue 6
Received: 22-Apr-2021
Published:
13-May-2021
, DOI: 10.37421/2161-0525.2021.11.633
Citation: Indo, Hiroko P, Hiromu Ito, Keiichi Nakagawa and
Hideyuki J. Majima. " Clarification of Mechanisms of Reactive Oxygen Species
Generation from Mitochondria”. J Environ Anal Toxicol 11(2021):633.
Copyright: © 2021 Indo HP, et al. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted
use, distribution and reproduction in any medium, provided the original author and source are credited.
It is known that reactive oxygen species (ROS) generated from mitochondria promote apoptosis [1]. Motoori et al. [2], Majima et al. [3] and Indo et al. [4] reported that X-ray’s irradiation causes ROS generation from mitochondria and leads to apoptosis. We also published evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage [5]. Recently, using human lung carcinoma cell lines A549 and H1299/wtp53, we demonstrated that 15.5 Gy monoenergetic X-ray irradiation of the K-shell resonance absorption peak of phosphorus results in increased mitochondrial ROS production [6]. The increased mitochondrial ROS generation due to X-irradiation suggests a common pathway with other types of irradiation that induce mitochondrial ROS generation [7,8]. However, the mechanisms by which ROS generate from mitochondria are unknown.
We published a paper titled “Translocation of HSP47 and generation of mitochondrial reactive oxygen species in human neuroblastoma SK-N-SH cells following electron and X-ray irradiation” [9]. In this study, we showed an increase in mitochondrial ROS generation following both 15 Gy X- and 10 Gy electron irradiation (Figure 1).

Figure 1: 1A: Detection of HPF fluorescence in the SK-N-SH cells irradiated with 0, 1, 3, 10 Gy and 15 Gy X-rays and with 0, 1, 3, and 10 Gy electron; 1B: Representative images of X-irradiated cells and electron irradiated cells in which ROS were visualized using HPF. A clear increase of fluorescence, and an increase of ROS was shown in 15 Gy X-irradiation and 3 and 10 Gy electron irradiation.
Intracellular localization of HSP47 (Figure 2), myristoylation (Figure 3), and farnesylation (Figure 4) following 15 Gy X- and 10 Gy electron irradiation was detected with immunohistochemistry. Cells were stained with Hoechst33342, MitoTracker Red, and ER staining dye to examine organelle localization. X-rays irradiation (15 Gy) and electron irradiation (10 Gy) induced intracellular HSP47 expression, myristoylation and farnesylation and HSP47 was all localized in mitochondria. (Figures 2-4).

Figure 2: 2A: Intracellular localization of HSP47 in 0 and 15 Gy X-ray-irradiated cells and in 0 and 10 Gy electron irradiated cells; 2B: Detected by immunohistochemistry Cells were also stained with Hoechst33342, MitoTracker Red, and ER staining dye to examine organelle localization. X-ray irradiation (15 Gy) and electron irradiation (10 Gy) elevated levels of HSP47 and HSP47 was localized in mitochondria.

Figure 3: 3A: Intracellular localization of myristoylation in 0 and 15 Gy X-ray-irradiated cells and in 0 and 10 Gy electron irradiated cells 3B: detected by immunohistochemistry. Cells were also stained with Hoechst33342, MitoTracker Red, and ER staining dye to examine organelle localization. X-ray irradiation (15 Gy) and electron irradiation (10 Gy) elevated levels of myristoylation and myristoylation was localized in mitochondria.

Figure 4: 4A: Intracellular localization of farnesylation in 0 and 15 Gy X-ray-irradiated cells and in 0 and 10 Gy electron irradiated cells 4B: Detected by immunohistochemistry. Cells were also stained with Hoechst33342, MitoTracker Red, and ER staining dye to examine organelle localization. X-ray irradiation (15 Gy) and electron irradiation (10 Gy) elevated levels of farnesylation and farnesylation was localized in mitochondria.
Transfection with HSP47 revealed that HSP47 is localized in the mitochondria and ER (Figure 5). 15 Gy X-ray’s irradiation and 10 Gy electron irradiation increased the levels of HSP47 (Figure 6), and the transfection of HSP47 increased mitochondrial ROS generation (Figure 7). These results suggest that HSP47 is located in the mitochondria where it promotes ROS production in human neuron-like SK-N-SH cells (Figure 8). Therefore, we clarified part of mechanism underlying ROS generation in mitochondria, for the first time.

Figure 5: 5A: Intracellular localization of HSP47 in HSP47-GFP-transfected cells. Double staining of HSP47 and MitoTracker Red shows that most of the HSP47 is localized in the mitochondria. 5B: Intracellular localization of HSP47 in HSP47-GFP-transfected cells. Double staining of HSP47 and ER shows that most of the HSP47 is localized in the ER.

Figure 6: HSP47 expression in mitochondria after 0 and 10 Gy electron and 15 Gy

Figure 7: 7A: Detection of ROS in HSP47 over-expressed cells, showing an increase in the mitochondrial ROS generation in HSP47 transfected cell 7B: Quantitative measurement of ROS using HPF dye in HSP47 transfected cells. Reprinted from ref. 9 with permission.

Figure 8: Schematic graph depicting the translocation of HSP47 from the ER into the mitochondria, resulting in an increase in mitochondrial ROS generation and further subsequent events such as lipid peroxidation and signal transduction. These phenomena may cause oxidative stress disease.
X-ray irradiations was analyzed using western blotting. HSP47 expression increased in the mitochondrial fraction following X-ray and electron irradiation. Mean ± S.E., Scheffe’s F test, *P<0.05, **P<0.01.
H.I. contributed to the whole analyses and discussion of the results. H.P.I. and H.J.M. contributed to the experimental planning and discussion of the results and in drafting the manuscript. K.N. contributed to all of these aspects of this study, including executing the final revisions. All authors approved the final manuscript prior to publication.
None.
No potential conflict of interest was disclosed.
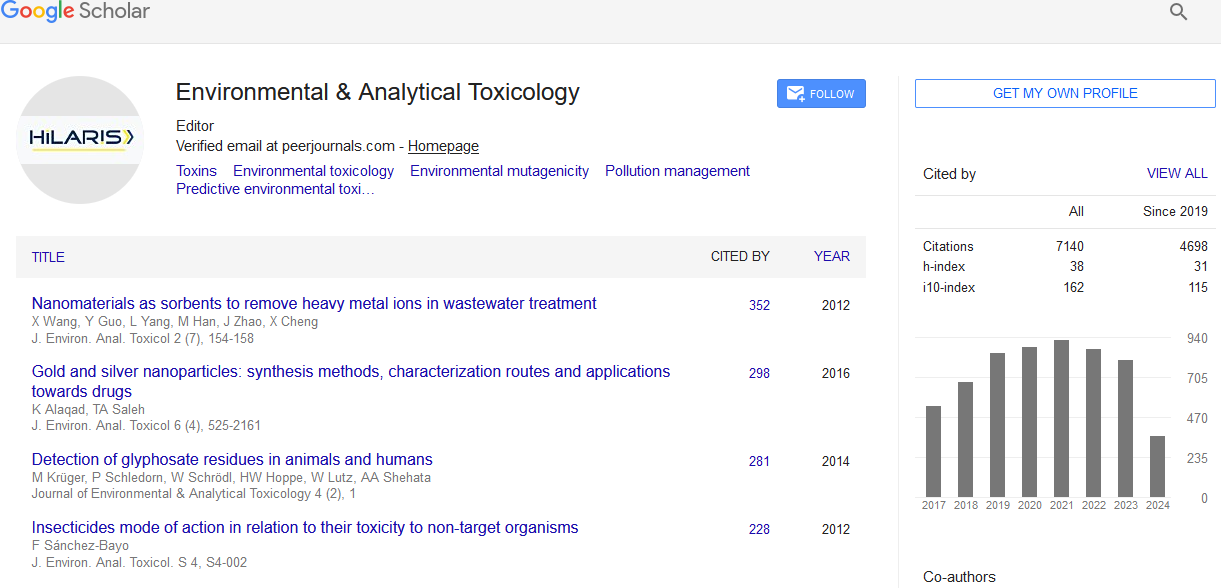
Environmental & Analytical Toxicology received 6818 citations as per Google Scholar report