Research Article - (2020) Volume 11, Issue 2
Received: 12-Dec-2019
Published:
17-Feb-2020
, DOI: 10.37421/2155-9619.2020.11.411
Citation: Bando M, Onoue Y, Tsunoyama Y, Suzuki K, Takanishi Y, et al. (2020) Comparison of the LQ and WAM Models on the Fractionation Effect in Radiation Therapy and Animal Experiments. J Nucl Med Radiat Ther 11: 410.
Copyright: © 2020 Bando M, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
The Linear Quadratic model (LQM) is an extended version of the linear non threshold (LNT) model, which has been used for quantitative description of radiation protection, radiation therapy and animal experiments for long time. However, the LQM encounters serious pitfalls, especially for the case of the fractionation irradiation. We propose the 'Whack a Mole' (WAM) model as a post LQM framework of the radiation and risk estimation, which includes cell exclusion effect. By introducing the time dependence, the WAM model explicitly includes not only the total dose but also the dose rate dependence. We show various important results of the WAM model for the dose rate dependence, the fractional irradiation effect, and the spontaneous decrease effect on mutation frequency. As an application of the WAM model, we apply the WAM model to the mega mouse experimental data taking into account the delay time of mating after irradiation.
LQ; WAM; Radiation therapy; Animal experiments; DDREF
The linear quadratic (LQ) model is commonly used for radiotherapy and/or radioprotection. It is a mechanistic, biologically based model with a few parameters and used for quantitative predictions of biological effects caused by radiation exposure and has been thought to estimate dose fractionation dependence for such cases as radiation protection, radiotherapy and animal experiments.
Historically the origin of the LQM can be traced back to X-ray mutagenesis of fruit fly discovered by Muller [1,2]. In the early period of study, scientists had understood that the radiation effects were calculated based on a simple physical process and its damage was irreversible and not preventable until a new concept appeared, which was developed in connection with dose rate and dose fractionation effects found in the mega mouse project by Russell and his collaborators [3-9]. This large scale project was proposed by Muller and Neel [10]. Those data published almost 50 years ago, however, are very important even at the present time, since the data replaced the previous concept that the radiation effects based on a simple physical interpretation, namely it is irreversible and not preventable, by a new one that the radiation damage can be reduced by repairing and cell exclusion mechanisms existing in the biological organism. Such an important message was not taken correctly because of the inconsistency between the data of fruit fly and mouse. Moreover a mathematical model of mutation frequency was proposed in terms of microscopic physical point of view, and the complete formulation was constructed by Lea [11]. This Lea's hit model elegantly reproduced the LNT behaviour, reproducing the established experimental results of the fruit fly. Since then the hit model has been thought to be an established model of radiation biology and has been the basis of radiation therapy and protection today.
However, the situation is quite different, when it is applied to the estimation of biological risk caused by a long term irradiation in a low dose rate circumstance as seen in high background radiation areas [12] or by the fractionation irradiation with long time intervals. Moreover there is a serious internal inconsistency in deriving the formula for the case of fractionated irradiation based on the LQM. This inconsistency comes from the lack of time dependence. Actually the more time a break is taken during the irradiation time, the larger the discrepancy between the results of fractionated and continuous irradiation cases becomes. This can be overcome by taking account of the temporal effects in the model. This is just the place where the dose rate dependence is important, the effect of which can be naturally accounted in the time dependent framework.
In this paper, we propose a model which aims at including explicitly the time dependence as well as the dose rate dependence by extending the LQ model, which has been most commonly used for quantitative predictions of dose fractionation dependence in radiotherapy. We shall show the origin of this discrepancy by demonstrating several examples.
This paper is written as follows. In method section, we make first a review of the LQ model together with the important indices, which have been used in radiological protection documents and papers referring the associated risk coefficients usually used. We discuss a serious internal inconsistency in the LQ model for the case of fractionation. We make a review of the WAM model with its characteristic features. We stress here that the WAM model is able to provide the spontaneous mutation frequency. The results of the WAM model on mutation frequencies for various dose rates while keeping the total doses same. We provide the mutation frequency as functions of the total dose and dose rate. We show important results of the WAM model on mutation frequencies for the fractionation case. We have presented the results of a reanalysis of the mega mouse experiment by taking into account the delay time of mating after irradiation. Finally the last section is devoted to the conclusion of this paper and discussions.
Rough sketch of LQM with the notion of DDREF
Let us start with a rough sketch of the essence of the LNT and LQ models, which are based on the standard Lea's hit model [11]. The hit model assumes, in a nutshell, that an important part in the cell (which nowadays is found to be the part located in the DNA sequence) is called "target" and if the radiation hits a target, a mutation and/or cell death occur. This hit process is expressed in terms of an independent probability event and the mutation frequency depends on the numbers of targets and hits. Thus the probability of hitting the target is determined by the total dose alone. This is a natural consequence of the standard calculation of probability of events happening independently. Therefore, regardless of the dose rate, the probability can be calculated depending only on the total dose, and independent of the interval of irradiation time. This is the essence of the target theory. We understand that it has become a common sense to express biological effects caused by radiation exposure in terms of "dose response relationship".
However, such a mathematical model cannot account for the dose rate effects, which were found to be needed to reproduce the mega mouse experimental data. We should construct a time dependent formalism because the damage repair exclusion balance is essential on the mutation frequency, which cannot be determined by the total dose D alone. This model indeed reproduces the fruit fly experiments done by Muller [1], which had been almost completely confirmed by the following many experiments [13-16]. This hit model was inconsistent with Russell's mega mouse data at that time, but the LNT hypothesis was adopted as a basic principle of radio protection following the wellestablished model in fruit fly experiments.
Biological Effects of Ionizing Radiation VII (BEIR VII) summarized the historical results and the well-established formulae [17]. Here, we make a quick review of the historical development using the mutation frequency as an example. From the mutation frequency F(D), let us de ne the excess mutation frequency E(D), by subtracting the initial mutation frequency,
E(D) = F(D = 0) (1)
Here, D is the accumulated total dose of artificial radiation, while F(D=0) corresponds the spontaneous (background) mutation frequency. In the LNT model,
ELNT D =α D (2)
with a parameter α being a coefficient of total dose D. This linear dependence is improved in the linear quadratic model (LQM) by including the quadratic term with coefficient β.
ELQM D =α D +βD2 (3)
in order to reproduce experimental data. Apparently in the above formula, dose rate effect is not taken into account explicitly. Instead the notion of DDREF (Dose and Dose Rate Effectiveness Factor) was introduced, which is a factor initially introduced by the International Commission on Radiological Protection (ICRP) to be used for the risk estimates of moderate to high dose and high dose rate data and for those of low dose and low dose rate data [18]. Note that the definition of DDREF changes with time (for example, Ruhm et al. reported on the long history from UNSCEAR 1962 (information from the atomic bomb survivors) to UNSCEAR 2013 (Comment on WHO report) [19]). Recently, a critical comment on the LNT hypothesis was made for the use in nuclear medicine and molecular imaging [20].
It seems, however, that the LQ model failed in making clear the difference between the dose and dose rate. Indeed in the expressions of ELNT and ELQM, there is no explicit dose rate d dependence and/or the concept of time t [21, 22]. This is the point we want to discuss in this paper. However, before that we should understand what people have been aiming at using the above LQM framework. One way of such approach can be seen from the discussion of Niwa, who assumed that the dose rate effects are clearly visible at the higher dose D region, while the linear term is independent of the dose rate, and the quadratic term is dose rate sensitive [19]. Then the biological effect for high dose rate exposures (H) and low dose rate exposures (L) can be described as, EH= α D+ β D2 and EL= α D. DDREF is defined as
![]() (4)
(4)
which is actually the same expression of the DDREF index as written in terms of the ratio of ELQM and ELNT: DDREF=ELQM=ELNT. This expression (4) is found also in the definition of biological effective dose (BED), which is frequently used in radiotherapy [23].
From the above discussion we understand that the notion of DDREF was indeed introduced not only to account for the dose rate effect but also to add a correction term to handle the data in the high dose region. Provably people might have naively imagined high D and low D correspond to high and low dose rates, respectively. However, if we encounter the situation of long term low dose rate exposure seen in some area of high background radiation, the correspondence is not justified, which we shall see later.
Moreover, the above complicated discussion added several notations, such as Dose Rate Effectiveness Factor (DREF), defined as the ratio of the effect at a given acute dose to that of a chronic exposure to the same total dose DREF=( α D+ βD2)=D. While the Low Dose Effectiveness Factor (LDEF) is defined as LDEF=1+( β / α )=D, which is used here and there in estimating radiation protection reports. However, they are almost the same, and may bring some confusion in scientific discussions. More confusing notion may be what is called "Committed Effective Dose" (CEO), which is defined as the time integral of the equivalent dose rate in a particular tissue or organ that will be received by an individual following intake of radioactive material into the body, where the integration time is as long as 50 years. ICRP actually tells how to calculate the CEO.
Let us make further an important comment on the notion of "fractionation effect" to be derived from the LQ model, which leads us serious pitfalls. The fractionated irradiation is utilized in the treatment of cancer in the radiation therapy or setting various experimental conditions for animal experiments. When the total dose of irradiation is divided into several smaller doses, people experienced that the radiation risk are lower for longer time interval. In the LQ model the excess dose in the fractionation treatment is formulated as
![]() (5)
(5)
indicating that the above procedure changes the coefficient ofD2 term, the contribution of the quadratic term becomes smaller for larger n. This equation 5 indicates that the mutation frequency decreases as the fractionation frequency n is increased.
However, we see an apparent inconsistency in the LQ model just by the following thought experiment. If we imagine the case where the time interval between irradiations is extremely short, namely almost instantaneous, there should be no difference between this fractionation procedure from the continuous irradiation. However, these two cases yield totally different predictions on the excess mutation frequency in the LQ model. For the continuous irradiation the excess radiation is E(D)= α D+ βD2. On the other hand, the excess mutation frequency in the n fractioned irradiation is written in equation 5. They are largely different. If we take n infinity as the limiting case
![]() (6)
(6)
We should be doing exactly the same irradiation in both cases; hence we ought to get the same results. But, they are totally different from each other. Why does it happen? The answer is simple. In the framework of LQM we have no information on the time dependence. Thus E depends only on the total dose D, which means that E stays constant so far as D does not change. However, we are aware of the fact that the actual data on mutation frequency change over time due to the decrease mechanism. Calculations of mutation frequency in clinical circumstance are getting important [21]. Taking account of such decrease effects, we have constructed a model, which will be presented in the next section.
We show a schematic diagram of the WAM model in Figure 1. Damage and repair processes occur frequently between normal and DNA damaged cells, and eventually a small fraction becomes mutated cell. Cell exclusion process as cell death happens from all the three cells expressed by the B term. The WAM model describes the transition to the mutated cell by the coefficient A and the cell exclusion process of the mutated cell by the coefficient B. In ordinary circumstance, all these processes occur constantly and hence we name this part as healthy state. Mutation in germ cell goes to hereditary disorder, and mutation in somatic cells, after escaping all the cell exclusion processes in various steps, may eventually grow to cancer, or leukaemia. These transitions are named genetic disease in Figure 1.

Figure 1. A schematic diagram of the WAM model, where normal cell, DNA damaged cell and mutated cell are interconnected. Genetic disease appears as a consequence of increase decrease competition expressed by the A and B terms in the WAM model. Even for the case without external stimuli (natural case), there is a increase decrease competition, and as a consequence mutation occurs for hereditary disorder, which we call a healthy state. Mutation in somatic cells, after escaping all the cell exclusion processes in various steps, may eventually grow to cancer, or leukemia.
The WAM model is able to describe the healthy state, where no artificial damages as radiation are applied to living bodies. It is natural to include the dose rate effect in the A and B terms in the WAM model to be able to describe the radiation effect on living bodies. In this model setting, we are able to know the status of the healthy state, which naturally leads to genetic disease state.
We have already stressed the importance of time dependence, because in a living system the competition of increase decrease mechanisms of the mutation frequency caused by transition to and exclusion of mutated cell are always in operation. The transition process should depend on the dose rate for the case of radiation. The exclusion process should have time scale of cell cycle in living body, and it should also depend on the dose rate. As a natural form of the event density, we take time dependent mutation frequency to explain the above statement more concretely. This should provide us with a powerful tool for further investigations towards cancer occurrence.
To see more clearly the mechanism of mutation, we express our dynamical equation in terms of a differential equation in time. We denote the mutation frequency F(t) changes by the competition of transition from normal DNA damaged cells and cell exclusion from mutated cells over the time.
![]() (7)
(7)
with the increase and decrease parameters A and B, representing the reaction rate from normal damaged cells to mutated cells (increase part of F(t)), and exclusion rate from the mutated cell (decrease part of F(t)). This equation is quite different from the Lea's hit model, which considers only the increase part.
Up to here the above equation is quite general. Only we have added the decrease term B to the hit model proposed by Lea [11]. By explicitly taking account of the dose rate dependence, we can estimate full effects of radiation in the form of fractionation or in different dose rate experiments to see clear difference between the LQ and WAM models. Although cells experience various kinds of processes, such as proliferation and reproduction from normal cells, as well as programmed cell death, and so on, we do not specify all the biological processes and just introduce effective reaction rates in the parameters A and B, including all the effects except the external stimulus terms coming from artificial radiation exposure.
Now we investigate the biological effects caused by radiation exposure. As a natural way we assume that the parameters, A and B, depend on the dose rate and they are written as A=a0+a1d and B=b0+b1d in equation 7. The most remarkable character of the WAM model comes from the additional term B, which represents all the cell exclusion effects. This is quite different from the LQM expressed in terms of the total dose D alone, and includes decreasing effects corresponding the second term with parameter B. Thus the mutation frequency decreases over time after the irradiation stops, while LQM predicts no change in the mutation frequency so far as the accumulated total dose D remains constant. Also we remark that the dose rate effect is automatically included through the parameters A and B. We can calculate mutation frequency caused by radiation exposure in a unified way for various cases as continuous, fractionated irradiation, or even post irradiation. The WAM model gives us a powerful tool to estimate the mutation frequency for various cases.
We comment here an important point on the LNT and LQ models, which are the hit model based on the target theory. These models are used independently both for the cases of mutation and cell death. However, it is important to point out that both the mutation and cell death happen simultaneously by a hit of radiation quanta on 'sensitive zone' in a cell. The sensitive zone for mutation is a particular locus in the DNA sequence, while the sensitive zone for cell death should be general loci. Hence, the probability of mutation should be much smaller than that of cell death. In the WAM model, these two processes are treated at the same time using the a1 and b1 terms, which are proportional to the dose rate d. We further comment that the cell exclusion process represented by the B term occurs on mutated cells, which implicitly assumes that the cell exclusion process occurs with the same probability on normal, DNA damaged and mutated cells.
In order to grasp the global structure of WAM by considering various cases, we first consider the case where d is constant during the irradiation interval, the solution of equation 7 can be obtained as,
![]() (8)
(8)
From the mutation frequency, we can write the excess mutation frequency.
![]() (9)
(9)
The behaviour of excess mutation frequency E(t) also depends on the initial condition F(t=0)=F(0). Its slope around the threshold can be even negative if F(0) is larger than A=B, which happens after irradiation.
Now, let us de ne several technical terms for later convenience. Spontaneous mutation frequency is defined as the mutation frequency coming from the natural effect even when there is no artificial irradiation (the case for d=0), which we denote Fs. Sometimes it is named as spontaneous mutation frequency or control or background term, which is obtained by taking d=0, with t infinity.
![]() (10)
(10)
It is important to note that the spontaneous mutation frequency Fs is obtained by the competition between the increase and decrease processes, where the spontaneous increase process is denoted by a0 and the decrease process is denoted by b0. Hence, even for the ordinary case without artificial irradiation, there is a competition of increase and decrease processes. In the healthy state, all the processes as cell damage, repair and cell exclusion are happening constantly. We see the consequence of the daily biological processes as the hereditary disorder for the germ cell and cancer and leukaemia for the somatic cell in the genetic disease state.
As we have already defined in equation 1, the excess effect should be defined by subtracting the mutation frequency F(t) by the spontaneous frequency Fs. For the continuous irradiation case with a constant dose rate d, the excess mutation frequency is
![]() (11)
(11)
Here, A=a0+a1d and B=b0+b1d. This expression can be rewritten in terms of the total dose D=dt, although E(D)(F(D) as well) include d dependent coefficients. By doing this we can get the relation of various kinds of effectiveness factors defined in method section DDREF [18, 19], which were derived from E(D). As for the case of fractionation irradiation, we replace the spontaneous frequency Fs by the value of the mutation frequency as F(0), which is the initial mutated value in the irradiation period with the dose rate d.
We introduce an effective dose-rate deff, which plays an important role in our later discussion.
A = a1 (deff + d) (12)
where deff=a0=a1. Here, deff is defined as equivalent dose rate, which induces the equivalent amount of spontaneous mutation frequency Fs. We note here that this equivalent dose rate deff causing the spontaneous mutation is much larger (almost 103 times larger) than the natural dose rate. This is extremely an important fact to note that there is a dominant source rather than the natural radiation, which induces mutation in living body.
We introduce then a critical time
tc = B−1 (13)
below which E(t) behaves linearly, and above which it deviates from the linear line and tends to a steady state. Namely tc measures the degree of resilience time. The critical time can be converted to the critical dose
Dc = dtc (14)
When the irradiation time t is larger than the critical time, i.e. t>B1=tc, we have
![]() (15)
(15)
For the case where the initial condition F(0)=Fs, the upper bound of the excess mutation frequency depends on the dose rate. For the lower dose rate, the terminal excess mutation frequency becomes lower, and tends to zero for the dose rate d goes to zero as it should be.
We make here a comparison of the LQ and WAM models. When the irradiation time t is short, t≪B-1=tc, and irradiation starts at t=0, the mutation frequency starts from the spontaneous mutation value Fs. This is just a situation, where the excess mutation frequency shows linear dependence on the total dose D, which is in agreement with the LNT hypothesis. Thus the excess mutation frequency becomes proportional to D with its slope independent of the dose rate d. It is compared with the LQM (3),
![]() (16)
(16)
We note here that this relation is obtained just by comparing the coefficients of the linear D term in the LQ model. It is not clear, however, whether in realistic cases LQM takes the value given by the above relation (16). This relation (16) is very important, because it is consistent with the data obtained by fruit fly. Then a question arises "what happens to the mega mouse experimental data?” which we shall see later numerically. As for the value β , it is impossible to find any correspondence of LQM to WAM. This is because WAM indicates that F tends to approach an asymptotic value in contrast to the increasing behaviour of LQM. If we dare to get the correspondence, we would have gotten β with a negative value. We shall discuss on this point in more detail in result section.
Mutation frequency in the WAM model
We have explained the characteristic features of the WAM model. We will see here realistic predictions in various cases. From here we take the parameters from those obtained by fitting the mega mouse data, where we did not consider the delay effect for mating after irradiation in Table 1 [24].
![]() (17)
(17)
| Parameters | ||
|---|---|---|
| a0 | (1/h) | 3.24 × 10−8 |
| a1 | (1/Gy) | 2.94 × 10−5 |
| b0 | (1/h) | 3.00 × 10−3 |
| b1 | (1/Gy) | 1.36 × 10−1 |
Our main purpose here is to grasp the essence of the cell exclusion effect and demonstrate what is needed for reasonable risk estimation caused by radiation. In any case let us demonstrate the characteristic features of the WAM model by taking typical examples. From equation 10, we can estimate the following numerical vales of the mutation frequencies. The spontaneous mutation frequency is Fs=1.08 × 10-5. We can calculate the critical time (13) as tc=(3.00 × 10-3+1.36 × 10-1d(Gy=h))-1, which depends on the dose rate d.
First let us see the global feature of the WAM predictions. We show in Figure 2 the mutation frequency as a function of the total dose D for the case of continuous irradiation with various dose rates d ranging from 0.01-1.0 Gy/h. Here we do not show figures with t dependence, since most of readers are accustomed to see only the D dependence.

Figure 2. Mutation frequency F (t) as a function of total dose D. Many curves represent F for various dose-rates ranging from 0.01 to 1 Gy/h. The dashed line is a linear extrapolation from the threshold region, where the threshold value of all the curves is Fs = 1.08 × 10-5.
We see an apparent dose rate effect, especially for larger D region. Let us first make comment on the global structure of high and low dose rate (HDR, LDR) cases. We recognize a clear difference between HDR and LDR. For the former case we recognize almost LNT behaviour for wide range of D, while for the LDR case it deviates from the LNT line largely at the critical dose Dc=dtc appearing at lower D. Especially for the LDR case, when the total dose becomes large, the mutation frequency is already at a saturation value. Even for the case of large dose rate, the asymptotic mutation frequency is F (t→∞, d=∞)=2.16 × 10-4. This saturation property can be confirmed in the following equation:
![]() (18)
(18)
If the dose rate is very small as less than 10 times of the natural radiation, the saturated value should be close to the spontaneous value. The details may be seen in the paper [25]. Also notice that around the threshold, the slope of F is common for all dose rates (indicated by dashed line in Figure 2), the value of which can be estimated from equation 16.
Therefore LNT is a good approximation so far as the total dose D is smaller than Dc, from where it gradually deviates from the LNT line and approaches their asymptotic values. We note that the critical total dose Dc depends on the dose rate d. The asymptotic values depend on the dose rate, and higher is the dose rate, the larger the asymptotic value is. Further, we can see that for cases of very high dose rate, the upper value itself tends to the terminal value: 2.16 × 10-4 as seen in equation 15. In this way we can confirm that the cell exclusion process during the time course is essential due to the balance of the increase decrease competition, which is demonstrated in terms of the total dose D in Figure 2.
We show the global feature of F(D;d) using Figure 3 by plotting F in (D;d) plane from a bird's eye view. This bird's eye view was first proposed by Strom [26] to understand the data obtained by the famous mega mouse project. This is obtained from Figure 2 with dose rate ranging from 0.001-0.1 Gy/h. Readers can understand at a glance how our predicted mutation frequency curves behave: If the readers put the observed animal data in this setting, then they can confirm what region each experimental data point is located. Examples may be seen in Figure 4 [27].

Figure 3. A bird’s eye view of mutation frequency F in the D; d plane. The parameters used are those of Wada et al. [25].

Figure 4. WAM prediction of the mutation frequency as a function of time for the case of fractionation and continuous irradiation. The upper solid curve corresponds a fractionated irradiation with 0.1 Gy/h for 100 h and 0 Gy/h for 100 h successively for six times. The upper dotted curve denotes a continuous irradiation with the dose rate 0.05 Gy/h. The middle solid curve corresponds a fractionated irradiation with 0.04 Gy/h for 50 h and 0 Gy/h for 50 h successively for 12 times. The middle dotted curve denotes a continuous irradiation with the dose rate 0.02 Gy/h. The lower solid curve corresponds a fractionated irradiation with 0.02 Gy/h for 100 h and 0 Gy/h for 100 h successively for 24 times. The lower dotted curve denotes a continuous irradiation with the dose rate 0.01 Gy/h.
Fractionation effect
As already mentioned in method section, usually in the LQM framework the fractionation is treated by changing the amount of fractionated dose, whereas in the WAM framework F can be equally applied to the case of t dependent dose rate. In this section we should explicitly express t dependence of F, because the fractionation operation is planned in terms of a time schedule as the starting time, ending time of irradiation and no irradiation intermission. This can be taken into account by using the time schedule in the WAM calculation. In this paper, we focus on fractionation irradiation in animal experiments. Let us consider a timetable with the same total dose D as shown in Figure 4. We see for the fractionation case F decreases during the time interval of no irradiation d=0 and increases during the time of irradiation with finite d. We see the average mutation frequency is taken over the whole time of irradiation schedule; it is similar to the value of F with continuous irradiation with the averaged dose rate.
Let us compare the above results with those of the case of LQM. In order to compare the two models, we have to multiply the parameters α and β .As for α we have already obtained in equation 16. As for β , the situation is somewhat complicated, because it also depends on D. We adopt this value from a review of the value of β determined from the animal studies. UNSCEAR (2012) decided that DDREF was around 2, namely
![]() (19)
(19)
which indicates ( β / α ) D ~ 1. Taking account of the fact that it was found that both acute and protracted exposures appeared to have approximately linear dose responses at the total doses between 0 and 1.5 Gy, we should adopt the value of D somewhere around 2Gy. Thus we here tentatively use α= 2.8 × 10−5 [1/Gy] and β=1 × 10-5 [1/Gy2], assuming the case D = 2Gy.
Up to here we have discussed the results of fractionated irradiation using the WAM model. We make further an important comment on the cell exclusion effect after irradiation is terminated. As for the report of the results of mega mouse experiments, they summarize their experiments as follows. The first observation is that the mutation frequency dropped dramatically by lowering the dose rate [10]. The second observation is that if the male mice were not immediately mated with female mice following irradiation, the mutation frequency also dropped precipitously. The longer the time between irradiation and mating, the lower the mutation frequency.
We have already confirmed the first observation in Figure 2. Let us examine the second observation. We can show some examples of such events. In Figure 5 we show various cases with different values of F(t=t0) at the time t0, when the irradiation is terminated and the time dependent values of F are all decreasing over time. Then the mating time from the time t0=1000h, with time interval Δt, the value of F decreases and finally goes to the control value. Namely all the mutation frequencies F are almost of the same value after 2000 hours (almost 12 weeks in this case) after irradiation stops. The simulation results of Figure 5 tell us that sometimes the experimental values of lower dose rates are not always smaller than those of higher dose rates, even if their experimental condition is made with the same total dose, the time interval maybe longer than the protracted exposure cases.

Figure 5. Mutation frequency F as a function of time t in hour. Many curves represent F for five dose-rates d = 0.02, 0.04, 0.06, 0.12 and 0.18 Gy/h in different time durations leading to the same total dose of D=18 Gy. The resulting mutation frequencies at the time t0 = 1000 h are different each other, and as the doserate is larger, the mutation frequency at t0 is larger. After the irradiation is terminated at this time, the mutation frequencies decrease with time t.
The above results teach us that in order to compare WAM with experimental data we really need to get exact time schedules of irradiation exposure. However, unfortunately, as for experiments in radiation biology, most experimental report did not mention about the details of time schedule, because at that time people believed that the mutation frequency depends only on the total dose and did not care about its time dependence, even in the mega mouse experiments done by Russell [9]. This is why we simply assumed that there is no time interval in irradiation experiments in our previous publications [25, 27, 28]. Recently we have investigated carefully the time schedules of the experiments and tried to reconsider the effect of the time schedule.
For example, our model predicted the mutation frequency caused by lower dose rate irradiation sometimes exceeds the one of higher dose rate exposure even their total doses are the same, and found it could have had happened [29].
Reanalysis of mega mouse data with WAM
In the era of mega mouse experiments, scientists did not consider the decrease process due to the cell exclusion expressed by the b0 term in WAM [7,8]. An example of the amount of the decrease effect is shown in Figure 5. Biologically we know that male mouse needs 7 weeks to make spermatozoon from spermatogonium, particularly for those involved in mouse experiments. Hence, the effect of irradiation appears in those off springs mated after 7 weeks from the time of irradiation. This duration time has been considered in the mega mouse experiments, although it is not mentioned in a clear manner in published papers [7,8]. As seen in Figure 5, the mutation frequency measured in off springs is decreased largely by about 30 times as compared to the mutation frequency just after irradiation. We ought to consider this effect in the WAM analysis for the mega mouse data. As stated in the end of the previous section, we do not have detailed information of the time schedule of irradiation and mating in the mega mouse experiments.
Considering this situation, we perform a parameter fitting in two steps. Since the spontaneous and the low dose data are almost completely determined irrespectively of irradiation time scheduling and sensitive to the parameters a0 and b0, we take the 5 points including the 3 points for spontaneous mutation n=1, 2, 3, 4, 5 in Figure 6. We × these parameters from this fitting procedure, and then take the 3 points of the high dose experiment (n=16, 17, 18) to × the high dose sensitive parameters a1 and b1 by maximizing the Poisson likelihood function. The parameters so obtained are
![]() (20)
(20)

Figure 6. The left figure is the mutation frequency as a function of total dose D for various dose rates. The black circles with error bars denote experimental results, while the red circles denote theoretical values. Several points for the same total dose are the results for different dose rates. The duration after irradiation up to the mating time is taken as 49 days in the WAM calculation. The right figure is the mutation frequency arranged in the order of the list of the paper of Russel and Kelly [8]. The dose rate independent parameters are fixed by using the lower 5 points numbered n = 1, 2, 3, 4, 5. Fixing a0 and b0, the dose rate dependent parameters are fixed by using the higher 3 points numbered n= 16, 17, 18.
These parameters are similar to the set of parameters of equation 17 except a1. The new parameter a1 is about 50 times more than that of the set of parameter (17), since the mutation frequencies just after the irradiation should be by far larger than those at the time of mating.
We provide the mutation frequency as a function of total dose D for various dose rates in the left figure of Figure 6. In the right figure we show the mutation frequency ordered as the list of a paper of Russel and Kelly [8]. As can be seen from these figures, the lower dose rate data points are reproduced almost perfectly, and the high dose rate data points are also reproduced nicely. As for other data points in between, the agreement between the experimental numbers and the theoretical numbers are reasonable, but some experimental points, especially n=11 and 12 (see the right figure of Figure 6), deviate largely from the theoretical values. We may have to consider the experimental schedules of the data points corresponding to experiments with intermediate dose rate regions more carefully on the fractionation procedure, while for acute dose rate experiments the exposure time interval is far less than 1 hour and we do not have to worry the time schedule. In any case, it is important to get more information on the mega mouse experiments and/or, if possible, new data points for variations of the experimental data points for detailed comparison of the theoretical prediction with experiment. Especially it is to be emphasized that the mutation frequencies change largely by the amount of the rest time in the radiation process due to the decrease effect.
We made a reanalysis of the mega mouse data using the LQ model also. Since the LQ model depends only on the total dose, the reproduction of each data point cannot be achieved and the theoretical results go through about the middle of data points with different dose rates for the same total dose. Hence, the LQ model fails to reproduce the two slopes obtained by Russell and Kelly in their original paper [8].
We demonstrated in the LQM framework there existed several unnatural aspects especially in the case of fractionation irradiation. This is due to the assumption that the mutation frequency F depends on the total dose alone and the effect caused by the "exclusion mechanism" was never taken into account. As a result, there is no time dependence in the expression of F in the LQM. On the other hand, the WAM model can overcome the above unnatural aspects by introducing the time dependence in F. As a consequence, the mutation frequency F explicitly depends on dose rate together with total dose. We showed the general features of the WAM model, where the mutation frequency depended largely on the dose rate even for the case of the same total dose. Especially in the case of fractionation irradiation, we compared the predictions of the WAM model and the LQ model. These two models showed totally different time profiles of the mutation frequency.
We formulated the increase and decrease effects represented by the A and B terms in the WAM model. The parameters of the WAM model had been extracted from experimental data of mutation frequencies in the hereditary effects from animal and plant experiments. We had already succeeded in reproducing dose rate dependences and the saturation behaviours of mutation frequencies for mega mouse and fruit fly data as well as many plants [25,27,28]. In this paper, we reanalysed the mega mouse data, since the WAM model made clear a large decrease effect on mutation frequency due to large time duration of mating after the irradiation [29]. We obtained a new parameter set for the mega mouse data, where the a1 parameter was found to be larger than the one of the previous parameter set, while other parameters were almost unchanged. The dose rate dependence was reproduced reasonably except for two points (n=11, 12 in Figure 6), which indicated the necessity of further attention on the time schedule of the mega mouse experiment. These parameters as well as those extracted from fruit fly and many plants are important for the study of mutation in cell beyond any species and biological evolution. As for the LQ model, there were very few studies on animal experiments, for which we were not able to reproduce the mega mouse data due to the fact that the dose rate dependence was not taken into account.
For the case of somatic cells, the pathways from mutation to cancer is not yet clarified since the process is thought to be induced via multi steps, involving growth and prevention, even if the first stage should be more or less related to mutation of cell [29]. However, even scientists in this field have not arrived at a final conclusion on the pathway from mutation to cancer in spite of quite a long research history. In light of this situation, the next target for the present research program should be the cancer therapy project, where a huge phenomenological data have been accumulated based on the LQ model together with the BED index. However, the parameters α and β obtained from the data, scatter from organ to organ largely [23,30,31] and depend on various stages of cancer growth [32] and the treatment plans have been decided in a complicated way using the parameters α and β above difficulties can surely overcome by our alternative model by taking account of the growth and decrement scenarios.
this paper, we mentioned a possibility of an application of the WAM model on the fractionation radiation therapy. We are now constructing a mathematical model based on the WAM model by treating cancer steps toward cancer as a black box, and the growth of cancer and the cell death effect due to irradiation should be taken into account. It is important to point out that the parameter α used in the cancer therapy in the LQ model is close to the value of the cell exclusion b1 in the WAM model. All the details of the application of the WAM model to the fractionation cancer therapy including the reanalysis of the ratio α / β tabulated in a review paper will be published in near future.
We are thankful to Prof. Yonekura for illuminating discussions on fractional irradiation for cancer therapy. This work was supported by the Ministry of Education, Science, ports and Culture, Grand in Aid for Scientific Research (B) 2016-2019 [16H04637, Takahiro Wada].
Dr. Suzuki reports nancial relationship from Medical corporation Midtown clinic, outside the submitted work. The other authors declare no competing interests.
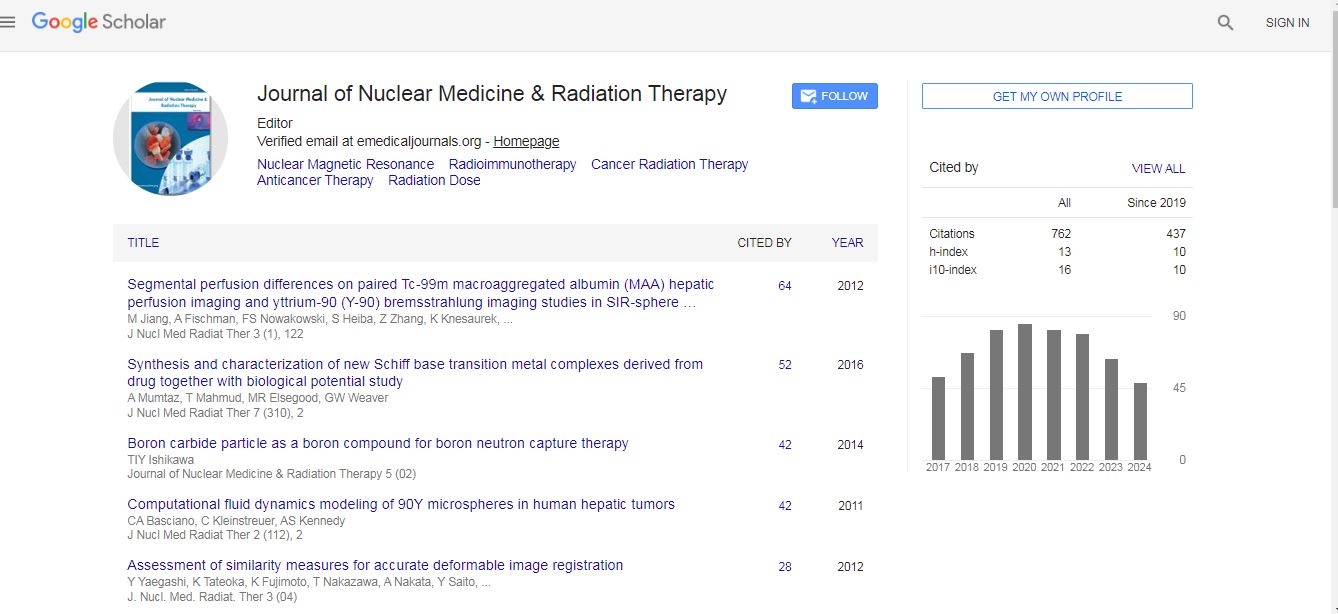
Nuclear Medicine & Radiation Therapy received 706 citations as per Google Scholar report