Brief Report - (2022) Volume 7, Issue 4
Received: 07-Jul-2022, Manuscript No. jib-22-77690;
Editor assigned: 09-Jul-2022, Pre QC No. P-77690;
Reviewed: 24-Jul-2022, QC No. Q-77690;
Revised: 29-Jul-2022, Manuscript No. R-77690;
Published:
05-Aug-2022
, DOI: 10.37421/2476-1966.2022.7.184
Citation: Michael, Mathieu. “Environmental Factors Alter Immunological Reaction with mTor.” J Immuno Biol 7(2022): 184.
Copyright: © 2022 Michael M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
The innate immune system plays a crucial role in maintaining tissue homeostasis and reacts swiftly to local or systemic perturbations caused by pathogenic or sterile shocks. To enable cell migration and proliferation, as well as the effective generation of cytokines and lipid mediators, this rapid response needs to be metabolically supported. This review focuses on how the innate immune system's effector responses are regulated and shaped by the mammalian target of rapamycin (mTOR). mTOR alters cellular metabolism and controls cell movement, antigen presentation, cytokine responses, translation, and polarization of macrophages. In order to control and optimize the inflammatory response, the mTOR network functions as an integrative rheostat that integrates cellular activation with the external environment and intracellular nutritional status.
In the 1970s, soil samples from Easter Island (locally known as Rapa Nui) were found to contain Rapamycin, the archetypal inhibitor of the mammalian target of rapamycin (mTOR), which was proven to have antifungal activity. It is now understood that mTOR has a variety of roles in mammalian cells and serves as a key regulator of cellular metabolism. As a result, targeting the mTOR pathway is used in immunosuppressive and anti-proliferative therapies for cancer and organ transplantation. Recent research has demonstrated the critical function of the mTOR complex 1 (mTORC1) and mTORC2 in controlling the populations of innate immune cells. Today, it is well understood that the mTOR signaling network acts as an integrative rheostat, coordinating a vast array of cellular and metabolic processes that influence immune effector responses. Each immune cell's reaction is customized and modified to meet its unique requirements based on the kind of cell, the target tissue, and the extracellular and intracellular nutritional status [1,2].
Innate immune cells that have been activated go through a major change in cell shape, move to new tissue locations, and release a lot of cytokines, chemokines, and lipid mediators like prostaglandins and leukotrienes. The mTORC1-mTORC2 network is responsible for shaping and controlling many of these metabolically demanding processes after cell activation. Growth factors, Toll-like receptor (TLR) ligands, cytokines, and other extracellular signals can all activate the mTORC1-mTORC2 network in innate immune cells. For instance, the growth factors granulocyte/macrophage colony-stimulating factor (GM-CSF) and FMS-related tyrosine kinase 3 ligand (FLT3L) induce mTORC1 activation in DCs and neutrophils3-5; TLR ligands activate mTORC1 and mTORC2 in human and mouse monocytes, macrophages, and DCs, as well as mTORC.
In contrast to resting fibroblasts and other primary cells, the mTOR pathway is remarkably inactive in these cells in vitro without these activating inputs. Numerous fundamental cellular activities, including translation and protein synthesis, cell growth, metabolism, and anabolic processes, are regulated by the activation of mTORC1 and mTORC2. For instance, following 4 hours of lipopolysaccharide (LPS) stimulation, murine macrophages and DCs experience a huge surge in protein synthesis that is predominantly reliant on the phosphatidylinositol 3-kinase (PI3K)-mTORC1 pathway. It's interesting to note that glycolysis and glucose influx don't appear to be factors in this early rise in protein synthesis. By directly phosphorylating the translation inhibitors eIF4E-binding protein 1 (4E-BP1) and 4E-BP2, which release eukaryotic translation initiation factor, mTORC1 controls both cap-dependent and capindependent translation of mRNAs [3,4].
mTOR activation in myeloid cells
A lot of ribosomal and translation-related proteins, as well as genes involved in metabolism, are encoded by mRNAs that contain a 5′ terminal oligopyrimidine tract (5′ TOP) or a pyrimidine-rich translational element (PRTE). mTORC1 is particularly effective at promoting the initiation of translation of these mRNAs. As additional well-studied targets of mTORC1-mediated phosphorylation, ribosomal protein S6 kinase 1 (S6K1; also known as RPS6KB1) and S6K2 (also known as RPS6KB2) trigger the phosphorylation and activation of S6 to promote protein translation. As a result, one important role of mTORC1 is to promote global protein synthesis upon innate cell activation, which further causes long-term metabolic reconfiguration of cells. In agreement with this, a recent study revealed that macrophages' adaptability to LPS stimulation results mostly via proteome remodelling [5].
The mammalian target of rapamycin (mTOR) network in innate immunity cells can be activated by a variety of inputs. Through their respective corresponding receptors, growth factors, Toll-like receptor (TLR) ligands, or cytokines activate mTOR complex 1 (mTORC1) and mTORC2. Different adaptors, including the small GTPase RAB8A in macrophages, are recruited to the receptor complex as a result of receptor activation, including class I phosphatidylinositol-3 kinases (PI3Ks)60. Phosphoinositide-dependent protein kinase 1 phosphorylates phosphatidylinositol-3,4,5-trisphosphate (PtdInsP3) on threonine 308 to recruit and activate the serine-threonine kinases AKT1, AKT2, and AKT3. PtdInsP32 is dephosphorylated by the enzyme phosphatase and tensin homologue (PTEN), which negatively regulates this activity. AKT is phosphorylated by mTORC2 on serine 473, which appears to be necessary for AKT's complete activation and substrate selectivity.
Additionally, in order to control crucial physiological processes including cytoskeleton remodeling, mTORC2 phosphorylates serum and glucocorticoidregulated kinase 1 (SGK1) and protein kinase C (PKC). Tuberous sclerosis 2 (TSC2) and Forkhead box O1 (FOXO1) are two of the key AKT effectors. When combined with TSC1, TSC2 produces a heterodimeric complex that blocks mTORC1. The small GTPase RAS homologue enriched in brain (RHEB) is phosphorylated at threonine 1462 (Thr1462) by AKT, which prevents TSC2 from activating mTORC1 on the lysosome through its GTPaseactivating protein (GAP) activity. The mitogen-activated protein kinases (MAPKs) p38 and COT activate mTORC1 in addition to the PI3K-AKT pathway through the phosphorylation of TSC2 by MK2 and ERK, respectively, in these pathways.
Metabolism, Innate Immunity, and mTor
It is now well accepted that for T cells to produce efficient defenses, appropriate metabolic reprogramming is necessary. There is growing evidence that innate immunity cells actively regulate metabolic functions in order to adapt and enhance their effector functions. Energy metabolism has been modified to accommodate these different effector activities' metabolic requirements and connect it to the availability of nutrients. Many fundamental metabolic activities are centrally regulated by the mTOR network. The production of nucleic acids, proteins, and lipids is often induced by the activation of mTORC1 through the hypoxia-inducible factor 1 (HIF1), peroxisome proliferator-activated receptor (PPAR), sterol regulatory element-binding proteins (SREBPs), and MYC [6].
It also promotes energy-producing procedures like glycolysis and mitochondrial respiration to supply the cellular energy and components needed for these reactions. By boosting the inactivation of class IIa histone deacetylases and activating AKT, mTORC2 also contributes to enhancing glycolytic metabolism. Forkhead box protein O1 (FOXO1) and FOXO3 are acetylated and rendered inactive as a result, activating MYC transcription.
We now have a plethora of knowledge on how the mTOR signaling pathway functions in innate immunity thanks to the recent years. A guiding paradigm suggests that the route, which further integrates the external and intracellular nutritional and energy status to direct and shape the effector response, is triggered by the activation of innate immune cells via pattern recognition or growth factor receptors. This happens in part as a result of the cell's metabolism being altered to supply fuel and building blocks for the cell's next immunological response. Additionally, mTOR modifies cell-specific central transcriptional networks such NF-B, STAT3, HIF1, and PPAR. A properly timed activation pattern of the mTOR network appears to be just as important as the fact that mTOR widely promotes many innate immune functions.
None.
The author shows no conflict of interest towards this manuscript.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
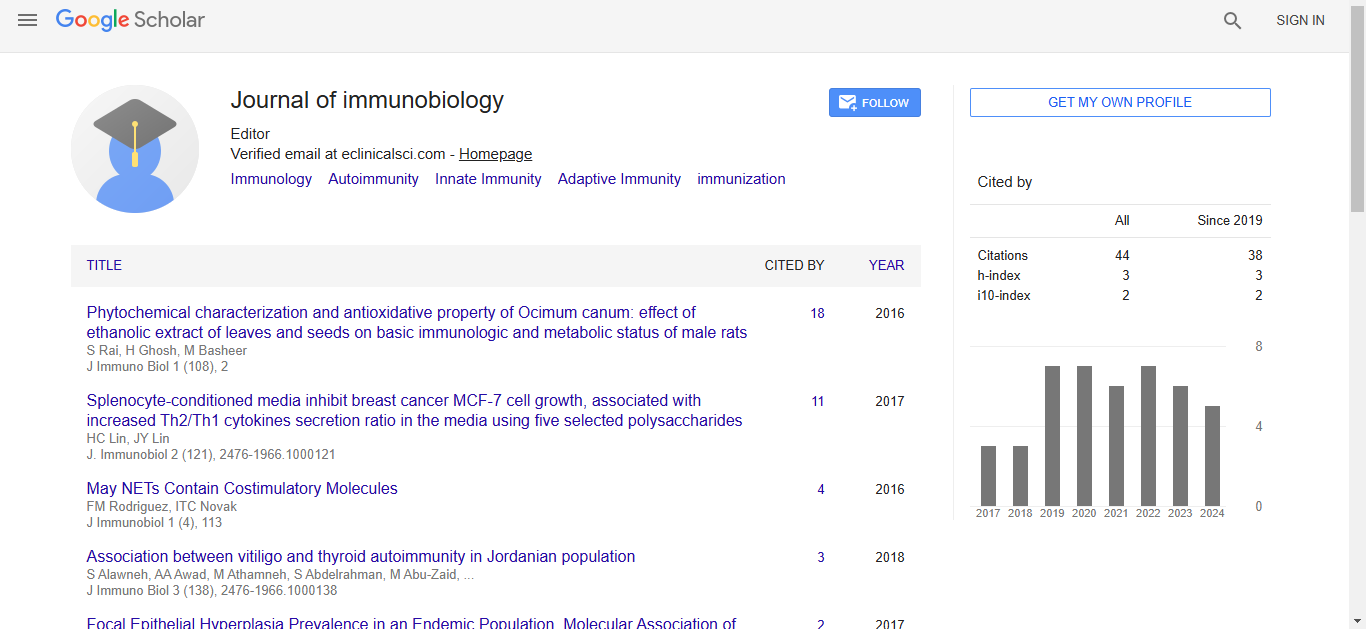
Journal of Immunobiology received 34 citations as per Google Scholar report