Research Article - (2023) Volume 13, Issue 4
Received: 20-Nov-2023, Manuscript No. JBL-23-120601;
Editor assigned: 24-Nov-2023, Pre QC No. JBL-23-120601;
Reviewed: 07-Dec-2023, QC No. JBL-23-120601;
Revised: 14-Dec-2023, Manuscript No. JBL-23-120601;
Published:
21-Dec-2023
, DOI: 10.37421/2165-7831.2023.13.314
Citation: Naderi, Majid, Maryam Emami, Mohammad Shojaei and Tahmine Davoodi, et al. " Evaluation of DNA Methylation and Genes Expression JAK2 and STAT3 Gene in Acute Lymphoblastic Leukemia." J Blood Lymph 13(2023):314.
Copyright: © 2023 Naderi M, et al. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Acute Lymphoblastic Leukemia (ALL) is the most prevalent malignancy among children. The primary therapeutic modality involves the administration of induction chemotherapy alone or in combination with diverse curative strategies. Existing evidence suggests that epigenetic mechanisms may serve as mediators of the influence of inherited genetic variations on phenotypic characteristics. Consequently, our investigation aimed to ascertain the potential role of DNA methylation in mediating the impact of genetic risk loci on childhood ALL. In mammals, the JAK/STAT pathway represents the principal signaling mechanism for a wide array of cytokines and growth factors. Activation of JAK induces cell proliferation, migration, differentiation, and apoptosis. A multitude of therapeutic interventions have been devised to modulate this signaling pathway, exhibiting varying degrees of efficacy and shortcomings. Pioneeringly, this study sheds light on the methylation status of JAK2 and STAT3, as well as the mRNA expression profiles, in ALL patients both prior to and following administration of the drug. We examined to determine whether there were any alterations in methylation and gene expression between the two genes during chemotherapy treatment.
This study, which took place from 2015 to 2017, utilized a case-control design. It included 50 blood samples from individuals recently diagnosed with Acute Lymphoblastic Leukemia (ALL) who had not yet received any chemotherapy drugs. After a two-month period of receiving the drug, these samples were retested. The study population consisted of 23 males and 27 females with a mean age of 7.52 ± 4.13. Additionally, 50 blood samples from healthy volunteers without any significant medical conditions were included in the study. This control group also consisted of 23 males and 27 females, with a mean age of 12.36 ± 5.63. All samples were stored at a temperature of -80°C until molecular analysis could be conducted. The methylation frequency of the JAK2 gene was found to be 35 (70%) in the blood sample taken from the newly diagnosed patient (referred to as sample1), 18 (36%) in the blood sample taken after the patient received chemotherapy (referred to as sample 2), and 3 (6%) in the blood samples from the healthy controls. The STAT3 gene exhibited a methylation rate of 54% (N=27) in sample1, 32% (N=16) in sample 2, and 4% (N=2) in the control group. A comparison between methylated and unmethylated samples indicated a significant disparity between the cases and controls in terms of JAK2 (OR1=36.55; 95%CI: 9.81 to 136.10, P < 0.0001) and STAT3 (OR1=28.17; 95% CI: 6.16 to 128.80, P<0.0001). Additionally, a notable distinction was observed between patients who underwent chemotherapy and the healthy individuals in relation to JAK2 (OR2=8.81; 95%CI: 2.39 to 32.40, P=0.0011) and STAT3 (OR2=11.29; 95%CI: 2.43 to 52.38, P=0.0020). In alternative terms, when we assessed the methylation status in patients subsequent to the administration of a chemotherapy drug in relation to their pre-treatment condition, a statistically significant finding was observed in JAK2 (OR3=4.14; 95%CI: 1.79 to 9.57, P=0.0009). However, no disparities were identified in STAT3 (OR3=2.49; 95%CI: 1.10 to 5). Within the framework of this investigation, we examined the disparity between the methylation of promoter DNA and the expression of genes within this pathway among patients who received the drug on the initial day, as well as the disparities with the control group. Chemotherapy drugs impeded the cell cycle and mitigated the adverse effects, particularly in the context of bone marrow metastasis, which was reliant on patient relapse in subsequent years.
ALL • Epigenetic • Methylation • Gene expression • JAK2 • STAT3
Acute Lymphoblastic Leukemia (ALL) is the most common cancer in children. The main treatment involves induction chemotherapy only or combination therapy with diverse curative approaches [1]. The cure success rate in Acute Lymphoblastic Leukemia (ALL) has grown steadily since the 1960s [2]. The five-year event-free survival rate is about 80% for children with ALL and almost 40% for adults [3]. If we include cases of relapsed ALL that respond well to so-called remission recovery treatment, the rates of cure (defined by the lack of evidence of sickness for at least 10 years) via modern medications are about 80% for children and 40% for adults [4]. Attempts to enhance the treatment rates through hematopoietic stem-cell transplantation have improved the outcome for some, but not all subtypes of ALL, suggesting that intensification of existing medications is unlikely to improve treatment rates substantially and will instead increase treatment-related death and the risk of life-threatening late sequelae such as second cancers [5]. Molecular analysis of the common genetic modifications in leukemic cells has contributed greatly to our understanding of the pathogenesis and prognosis of ALL. While the frequency of particular genetic subtypes differs in children and adults, the common mechanisms underlying the induction of all types are similar [6]. They include aberrant expression of proto-oncogenes, chromosomal translocations that produce fusion genes encoding active kinases and modified transcription factors, as well as hyperdiploid including more than 50 chromosomes. Evidence suggests that epigenetic mechanisms might mediate the effect of inherited genetic variations on phenotypes [7]. Now, we investigated whether DNA methylation mediates the effect of genetic risk loci for childhood ALL [8]. The Janus Kinase /Signal Transducers and Activators of Transcription (JAK/STAT) pathway are one of a handful of pleiotropic cascades employed to transduce a large number of signals for development and hemostasis in animals, from humans to flies (Figure 1) [9]. In mammals, the JAK/STAT pathway is the major signaling mechanism for a wide range of cytokines and growth factors [10]. JAK activation stimulates cell proliferation, cell migration, differentiation, and apoptosis. These cellular events are significant to hematopoiesis, immune development, mammary gland development and lactation, adipogenesis, sexually dimorphic growth, and other processes [11]. JAK/STAT signaling is a key factor in cancer progression, either as a tumor intrinsic driver of a metastasis/cancer growth factor or as a modulator of immune surveillance [12]. Activating mutations are reported in cytokine receptors upstream of JAK/STAT; for example, IL7RA, found in the extracellular and transmembrane regions in Acute Lymphoblastic Leukemia (ALL) [13]. Numerous therapeutics have been developed to moderate this signaling pathway, with varying degrees of success and failures. The difference in drug targets and efficacy aside, our emerging understanding of the significance of JAK/STAT signaling in both immune regulation and cancer progression has sparked significant research efforts [14]. Despite the recognition of the JAK/STAT pathway many decades ago, the knowledge of the details involved in this signaling cascade in cancer is still in its infancy. Epigenetic modifications such as DNA methylation of CpG islands in promoter regions are the major cause of tumor suppressor gene silencing and can result in tumor development. To our knowledge for the first time, this study highlights the status of JAK2 and STAT3 methylation and mRNA expression profiles in ALL patients before receiving the drug, and thereafter. We check if there are any changes in methylation and gene expression between the two genes in chemotherapy or not (Figure 1).

Figure 1. JAK/STAT pathway signaling. Created with BioRender.com.
Subject
This case-control study was performed from 2015 to 2017 comprising 50 (recently diagnosed without any chemotherapy drug) ALL blood samples which were again tested after receiving the drug following two months (23 males and 27 females with a mean age of 7.52 ± 4.13) along with 50 healthy blood samples of volunteers who had no special medical problems (23 males and 27 females with a mean age of (12.36 ± 5.63). The samples were frozen at -80°C until molecular analysis. These samples were collected from Imam-Ali Hospital. Methods in this study were approved by the Ethical Board at the Zahedan University of Medical Sciences. Informed consent was taken from all participant’s legal guardians [15].
DNA extraction and modification
Genomic DNA was extracted from the fresh blood vein sample via phenol chloroform isoamyl alcohol extraction protocol. Next, 1-2 μg of isolated DNA was diluted in 20 μg of distilled water and used for bisulfite treatment by EpiTect Fast Bisulfite Conversion Kits (Qiagen), which converted unmethylated cytosine to uracil and left methylated cytosine unaltered. According to the manufacturer’s protocol (Qiagen), the treated DNA should be diluted in 20 μL of distilled water and kept at -20°C for use in further experiments.
Methylation-Specific PCR (MSP)
To carry out the MSP analysis, promoters of the genes were recognized in online data analysis after which the chosen sequences were used to design methylated and unmethylated primers by MethPrime online software. Our choice for the site of methylated and unmethylated was the same with one received from the software. Hotstar Taq Master Mix Kit- Qiagen was used for each PCR reaction. Each PCR reaction contained 1 μL of modified DNA and 0.5 μL of each primer, dissolved in the vial of Hotstar Taq Master Mix Kit- Qiagen reaching the final volume of 20 μL with distilled water.
The MSP amplification was set as follows: 95°C for 5 min, followed by 30 cycles (95°C for 40 s, the annealing temperature for JAK2: M=58, U=57; STAT3: M=59, U=55 for 40 s and extension at 72°C for 40 s). The final incubation was completed at 72°C for 10 min. The designed primers are listed in Table 1. PCR products were detected via electrophoresis in 2% agarose gel, 80-100 volts for one hour until being well separated. Representative primer of methylation-specific PCR analysis of JAK2 (a) and STAT3 (b). M, methylated; U, unmethylated (Table 1).
| Genes | Sequences (5Ø?-3Ø? ) | Annealing temperature ( °C ) | Product size (bp) |
|---|---|---|---|
| JAK2 M | F:TGGTAGTCGGGAAGTTCGTTA R:AATAAAAATAAAATAAAACGCCC |
58 | 138 |
| JAK2 U | F:GGTAGTTGGGAAGTTTGTTA R:AAAAATAAAAATAAAATAAAACACCC |
57 | 141 |
| STAT3 M | F:TGTCGGAATAGTTAGTATAGGGGC R:CCAATACGTATACGATACACAACCG |
59 | 184 |
| STAT3 U | F:GTTGGAATAGTTAGTATATAGGGGTG R:TCCCAATACATATACAATACAATACAACCAAA |
55 | 186 |
RNA extraction and cDNA synthesis: Total RNA was extracted from blood samples by the Total RNA Extraction Kit-Pars Tous. Easy cDNA Synthesis Kit-pars Tous was employed to reverse-transcribe 1 mg of RNA to cDNA at a final volume of 20 μL.
mRNA quantification by real-time PCR: Real-time PCR was performed via SYBR green in ABI 5700 sequence detection system (Applied Biosystems). We compared the mRNA expression in ALL samples related to the control sample. β-actin was used as an internal standard. PCR Efficiencies (E) were calculated for all used primers from the given slopes of standard curves, produced from serial dilutions of positive controls, according to the following equation: E=2(−ΔΔCT). The designed primers for expression analysis are reported in Table 2.
| Genes | Sequences(5Ø?-3Ø? ) | Annealing temperature ( °C ) | Product size (bp) |
|---|---|---|---|
| JAK2 | F: CCCTCCATTTCTGTCATC R:AAGCAGGCAACAGGAACAAG |
59 | 594 |
| STAT3 | F:GACTCTCAATCCAAGGGGC R: CCTCTGCCGGAGAAACAG |
60 | 451 |
| β-actin | F:AGAGCTACGAGCTGCCTGAC R:AGCACTGTGTTGGCGTACAG |
65 | 524 |
Statistical analysis
The results of JAK2 and STAT3 genes methylation on the risk of ALL were identified by estimating Odds Ratios (OR) and 95% confidence intervals (95% CI), using logistic regression. To prevent bias in estimating OR, we calculated confidence intervals by three methods including exact, Cornfield, and Woolf. The Stata SE (version 13.1) was utilized for statistical analyses. The One way ANOVA test was used to compare expression data between groups. The statistical significance was set at P ≤ 0.05.
The methylation frequency of the JAK2 gene was 35 (70%) for sample 1 (newly diagnosed patient), 18 (36%) for sample 2 (patients after receiving chemotherapy drug) and 3 (6%) for healthy controls. STAT3 gene showed 54% (N= 27) methylation in sample 1, 32% (N=16) in sample 2 and 4% (N=2) in the control group. A comparison of methylated versus unmethylated showed a significant difference between cases and controls in JAK2 (OR1=36.55; 95%CI: 9.81 to 136.10, P<0.0001) and STAT3 (OR1=28.17; 95% CI: 6.16 to 128.80, P<0.0001). Also, there was a significant difference between patients after receiving chemotherapy drugs and the healthy one in JAK2 (OR2=8.81; 95% CI: 2.39 to 32.40, P=0.0011) and STAT3 (OR2=11.29; 95% CI: 2.43 to 52.38, P=0.0020). In other words, when we evaluated the status of methylation in patients after receiving chemotherapy drug with their pre-treatment state, it was significant in JAK2 (OR3=4.14; 95%CI: 1.79 to 9.57, P=0.0009) but no differences were found in STAT3 (OR3=2.49; 95%CI: 1.10to 5) (Table 3).
| Gene | Methylation status | ALL samples 1 N=50 |
ALL samples2 N=50 |
Normal samples N=50 |
OR1 | OR2 | OR3 | 95% CI | P-value |
|---|---|---|---|---|---|---|---|---|---|
| JAK2 | M U |
35 15 |
18 32 |
3 47 |
36.55 | 8.81 | 4.14 | 1=9.81 to136.10 2=2.39 to 32.40 3=1.79 to 9.57 |
1=P < 0.0001 2=P=0.0011 3=P=0.0009 |
| STAT3 | M U |
27 23 |
16 34 |
2 48 |
28.17 | 11.29 | 2.49 | 1=6.16 to 128.80 2=2.43 to 52.38 3=1.10to 5.62 |
1=P<0.0001 2=P=0.0020 3=P=0.0277 |
Note: Sample 1: Newly diagnoised patients who received no chemotherapy drug, sample 2: patients in sample 1 after receiving chemotherapy drugs, M: Methylated, M/U: the samples showed both methylated and unmethylated at the same time, U: Unmethylated, OR: Odd Ratio (1=sample 1 and normal, 2=sample 2 and normal, 3=sample 1, sample 2).
Decreased expression in the case group of both candidate genes was detected (29.21 ± 1.36 in case versus 22.47 ± 2.33 for controls in JAK2 and 26.39 ± 2.33 in cases versus 22.56 ± 2.55 controls in STAT3). There were differences in expression between patients after receiving the drug and healthy cases (24.90 ± 2.74 in case versus 22.47 ± 2.33 for controls in JAK2 and 24.84 ± 2.66 in cases versus 22.56 ± 2.55 controls in STAT3). Comparison of the mean between cases and controls revealed a statistically significant difference in both genes (P<0.05) (Table 4).
| Genes | N=15 | Mean ± SD | P value |
|---|---|---|---|
| JAK2 | Sample 1 Sample 2 Control |
29.21 ± 1.36 24.90 ± 2.74 22.47 ± 2.33 |
0.000* |
| STAT3 | Sample 1 Sample 2 Control |
26.39 ± 2.33 24.84 ± 2.66 22.56 ± 2.55 |
0.001* |
Note: Sample 1: Patients with new detection; sample 2: Pateints after getting chemotherapy drug, control: Healthy cases,*significant.
The principal result is that the cancer genome is limited and predictable, but is extremely complicated. Recent studies in epigenetics have suggested that human cancer cell harbor universal epigenetic abnormalities in addition to various genetic modifications and is a key to initiation of events in different types of cancer. Epigenetic abnormalities affect cancer chemotherapy response by modifying the expression of key genes involved in the absorption, metabolism, and excretion of drugs or those correlated with the progression or severity of cancers. Typically, the dosages reported in chemotherapy are lower than the actual dosage.
DNA methylation provides an epigenetic technique for controlling gene expression, so it is a good target for chemotherapy drugs. Studies have shown that JAK/STAT signaling pathway plays an important role in cell proliferation in cancer cells. Gao, et al. showed that silencing of LPCAT1 (an enzyme involved in phosphatidylcholine metabolism) would strongly reduce IL-6, p-Janus Kinase2 (JAK2), and p-STAT3 expression levels in cervical cancer cells [16]. Zhang et al. reported that JAK2/STAT3 pathway may represent a promising target for therapeutic intervention in osteosarcoma [17]. Also, Liang Chen, et al. showed that JAK2/STAT3 pathway repressing assay found that inhibition of the JAK2/STAT3 pathway significantly reduced EMT, cell migration, and stem cell sphere formation in prostate cancer [18]. Meanwhile, Xiao, et al. showed that treatment with MEX3C-specific antisense oligonucleotide inhibited JAK2/STAT3 pathway activation, suppressing hepatocellular carcinoma migration in vitro and metastasis in vivo [19]. Combination treatment with paclitaxel (a chemotherapy medication) and JAK2/STAT3 inhibition prevented development of the mesenchymal chemo-resistant subpopulation in breast cancer [20]. These results provide a mechanistic rationale for combining chemotherapy with inhibition of JAK2/STAT3 signaling as a more helpful therapeutic strategy in cancer. The study of the molecular mechanism showed that the JAK2/STAT3 pathway was involved in the DCZ0858-mediated antitumor effects and that JAK2 was the key target for DCZ0858 treatment. Knock down of JAK2 partially destabilized the DCZ0858 effect in Diffuse Large B-cell Lymphoma (DLBCL) cells, and JAK2 overexpression strengthened the effect of DCZ0858 in these cells [21]. According to the observations, ALDH1A1 (a detoxifying enzyme, that is responsible for chemoresistance in various tumors) induced resistance to CHOP (cyclophosphamide, doxorubicin, vincristine, and prednisone) during activation of the JAK2/STAT3 pathway in DLBCL, and research focus on it provides a potential strategic approach for reversing CHOP resistance [22].
In this study, we evaluated the difference between promoter DNA methylation and gene expression of two key genes of this pathway among the patients who received the drug on the first day as well as differences with the control group. Chemotherapy drugs inhibited the cell cycle and reduced the side effects, especially in bone marrow metastasis which depended on relapse in patients in the next years.
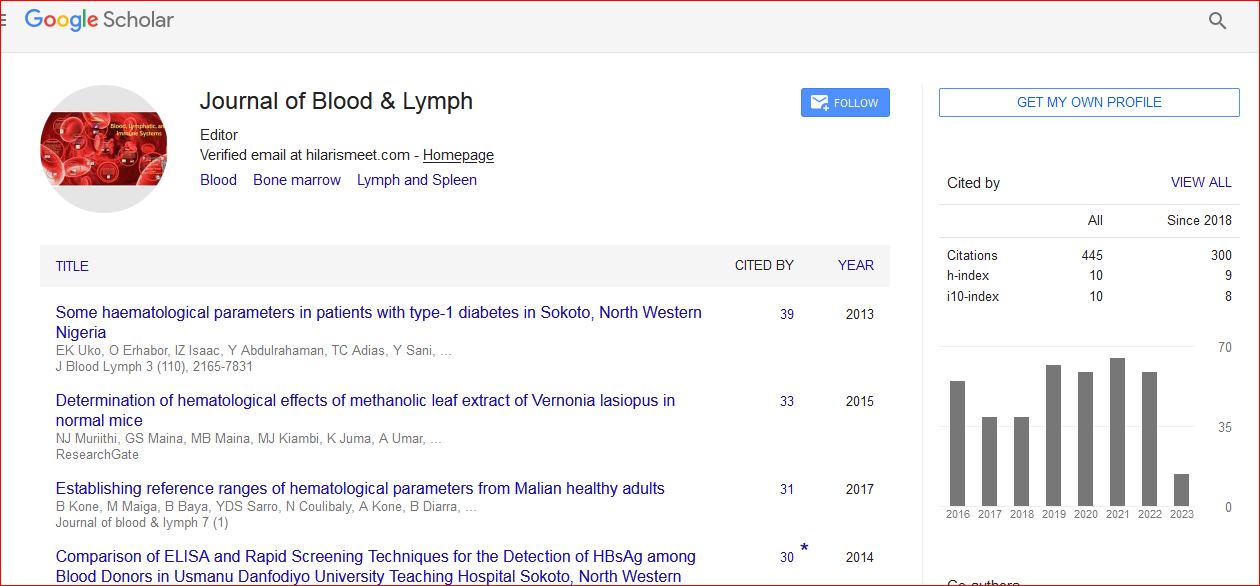
Journal of Blood & Lymph received 443 citations as per Google Scholar report