Review - (2022) Volume 10, Issue 10
Received: 02-Oct-2022, Manuscript No. jbes-22-82170;
Editor assigned: 04-Oct-2022, Pre QC No. P-82170;
Reviewed: 15-Oct-2022, QC No. Q-82170;
Revised: 20-Oct-2022, Manuscript No. R-82170;
Published:
26-Oct-2022
, DOI: 10.37421/2332-2543.2022.10.451
Citation: Taveira, German. “Expected Benefits of Plant Microbiome Engineering for Better Crop Growth and Resilience.” J Biodivers Endanger Species 10 (2022): 451.
Copyright: © 2022 Taveira G. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Microbiomes that are found in plants can help plants develop faster or manage diseases. A consortium of plant growth-promoting rhizobacteria (PGPR) can be inoculated into the microbiome to change it, which can improve plant growth and protect it from biotic and abiotic challenges. An innovative biotechnological method for increasing agricultural yields and resilience involves manipulating the plant holobiont through microbiome engineering. Direct methods of microbiome engineering include inoculation with particular probiotic microbes, artificial microbial consortia, and microbiome breeding and transplantation. Indirect methods involve the use of soil amendments or selective substrates. We discuss the benefits and potential integration of microbiome services into conventional agricultural methods as well as the knowledge gaps that need to be filled before these methods can be applied commercially in the field. Enhancing plant functions, including those related to biotic and abiotic stressors, plant fitness, and productivity, is the primary objective of microbiome engineering.
Plant microbiome • Crop growth • Plant holobiont
As a new paradigm for removing numerous microbiome-based agromanagement barriers, many ecological, biochemical, and molecularbased techniques have emerged. Additionally, multidisciplinary techniques offer a framework for prediction in order to create a plant-microbiome that is dependable and sustainable for stress physiology, nutrient recycling, and highyielding disease-resistant genotypes. Over the millennia, improvements in science have made microbial coevolution and diversification significant forces in shaping and carving every area of nature that is accessible. One of the key characteristics of plants becoming terrestrial is the interaction of taxonomically diverse microbial communities with various ecosystem components. The "holobiont" framework is thus shaped by this multi-organismal assemblage and its synergistic interactions with the host. The term "plant microbiome" refers to the ecologically diverse microbial populations that colonise plants in an integrated and dynamic manner [1,2].
Plant-microbiome interactions are influenced and manipulated by soil amendments, which are both organic and inorganic additions to the soil or agricultural techniques that farmers use to maximise output. The parts that follow give some illustrations. Lime, vermiculite, perlite, and sand are examples of inorganic supplements that have been used to lessen the effects of soil acidity and salinity on agricultural plants. Even in N-deficient soils, they can result in a larger, more active microbiome as well as an increase in plant biomass brought on by an increase in root exudates. Crop yield increased by 30% when inorganic and organic amendments were combined, but only by 8% when organic fertilisers were used alone. Throughout the plant's life cycle, the microbiota can live permanently in, on, and around various tissues. The plant's diverse microbiome includes bacteria, fungus, protozoa, viruses, and archaea. Host-microbe interactions are now better understood from an ecoevolutionary and functional standpoint thanks to research on their multitrophic interactions. The major microecosystems of the rhizosphere, endosphere, and phyllosphere are where bidirectional chemical communication directly affects plant development, physiology, and systemic defences and indirectly results in the production of root exudates and other metabolites that serve as sources of nutrients and signals for modifying microbial composition. Host genotypic characteristics, developmental stage, soil characteristics, and environmental factors balance the microbiome's structural and functional dynamics. Numerous adaptive benefits are provided to plant growth and development by this symbiotic interactome [3].
The multitrophic connections found in the mysterious plant microbiome have been considerably revealed thanks to the development of novel highthroughput community analysis methodologies, next-generation sequencing technologies, and meta-"omics" tools. These high-throughput technologies have made it possible to control helpful bacteria in plants. In this case, microbiome engineering might be a different approach to comprehend, control, and create corresponding technologies for microbial communities essential to the health and productivity of plants. The current analysis promotes a number of useful strategies for manipulating plant-microbial mutualism to improve plant performance and agricultural productivity. In microcosm tests, soil inoculants made of biofertilizers and biostimulants have been proven to be helpful for increasing plant productivity and defence against infections more broadly. However, a small number of recognised microbial strains that have been identified from a few plant species are present in the studied soil inoculants. Since these strains were developed in soil subjected to intense agricultural practises, it is reasonable to wonder whether the benefits they offer are limited to these particular circumstances. In this regard, studied the impact of agricultural approaches such as organic and conventional management and discovered that endophyte prevalence and variety were increased in 32 crops, including maize, tomato, melon, and pepper, by organic farming [4].
Plant microbiome
Years of study have indicated that a swarm of bacteria spontaneously colonises plants. Recognition changed from being given to lone entities to being given to communities. These bacteria are gathered in micropockets that are either on the plant's surface (phyllosphere), inside the plant (endosphere), or outside the plant (rhizosphere). These three major microecosystems are linked and have chemical communication zones. Rhizosphere refers to the crucial region where plant roots interact closely with the biotic and physicochemical elements of the soil, supporting a wide variety of microbial communities. The primary factors encouraging microbial community abundance and metabolic synchronisation are plant-root exudation and microbial substrate consumption characteristics. Complex microbial communities with a non-harmful nature that live and persist within plant tissues are what make up the plant endosphere. As a single habitat, it benefits more than the rhizosphere and is unaffected by microenvironment fluctuation and dynamics. Another microhabitat is the phyllosphere, which is made up of plant parts that are above ground and creates an oligotrophic environment for supporting a variety of microbial communities [5].
Evolutionary studies have supported the critical function of microbial phylotypes in ecosystems. These changing microenvironments serve as a critical link between top-down and bottom-up microbial diversity. By fusing the Green Revolution and agricultural sustainability, collective microbial habitats present an additional benefit. Agroecological systems can benefit from the beneficial microbes in the microhabitat in a variety of ways, including nutrient solubilization, phytohormone production, nitrogen fixation, bioremediation, improved tolerance to biotic and abiotic stresses, altered plant phenology, modified morphological and size-related traits, and ultimately played a significant role in mediating plant responses to climate change and variation. The content and activities of the associated microhabitat are influenced by molecular signalling and trophic interactions between the plants.
None.
None.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
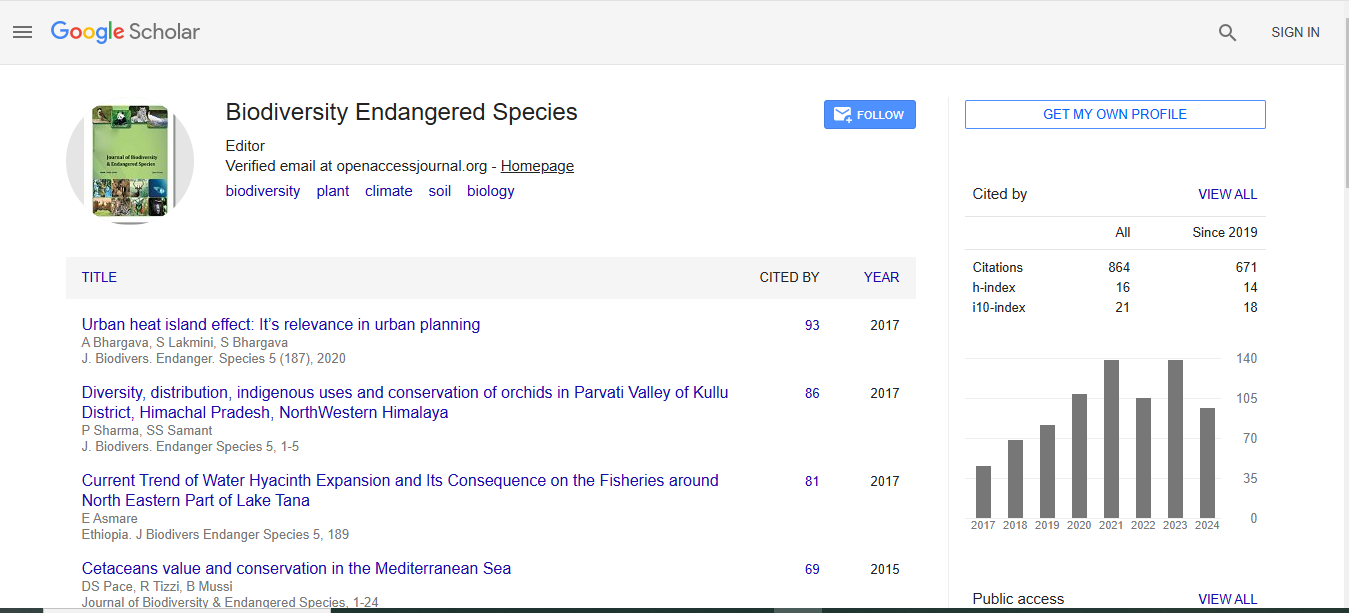
Journal of Biodiversity & Endangered Species received 624 citations as per Google Scholar report