Opinion - (2022) Volume 8, Issue 6
Received: 01-Nov-2022, Manuscript No. jfim-23-85460;
Editor assigned: 03-Nov-2022, Pre QC No. P-85460;
Reviewed: 15-Nov-2022, QC No. Q-85460;
Revised: 21-Nov-2022, Manuscript No. R-85460;
Published:
28-Nov-2022
, DOI: 10.37421/2572-4134.2022.8.261
Citation: Hammons, Emily. “Genome-Scale Metabolic Models for Food Microbiology: Applications and Drawbacks.” J Food Ind Microbiol 8 (2022): 261.
Copyright: © 2022 Hammons E. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Food fermentation began with uncontrolled process conditions and spontaneous fermentation by natural microbial consortia. The production became more regulated with the introduction of backsloping and knowledge of microbes. At an industrial scale, people are understanding, optimizing, and designing the conditions and microbes of the fermentation process. The shelf life and safety of food products, specific biotransformations, and the enrichment of food with desired properties are frequently linked to the goals of food microbiology. In order to stop harmful microbes from growing, food preservation requires a rapid acidification process. The preparation of indigestible food sources like coffee or cacao and the use of alternative dairy products like oat milk are examples of biotransformations. The improvement of food incorporates flavor compounds, organoleptic properties, the increment of nourishment worth or strains with probiotic properties.
The methods for achieving these goals have also improved. The systematic collection and classification of strains marked the beginning of advancements and continues to this day. The phenotyping zeroed in on morphology, development or fermentation properties. Today, it includes whole genome sequences, and multi-omics tools routinely monitor strain prevalence and activity in complex food matrices. It has remained challenging to bridge the gap between mere description and more mechanistic comprehension despite advancements in technology and data. Here, metabolic modeling has a lot of potential, especially when done with Genome Scale Metabolic Models (GSMM). As a collection of associations between genes, proteins, and metabolic reactions, these GSMMs organize an organism's metabolism. As a result, these models can predict metabolic capabilities based on genomes and provide a metabolic context for integrating data from cell physiology and genomics. There are numerous tools available to assist with each stage of the process.
The key goals of food microbiology have been addressed through the use of metabolic models, which we summarize here. We will emphasize open questions and the difficulties and restrictions throughout. We also include relevant and we believe promising examples of modeling outside of the food industry; This includes the most recent advancements in microbial community modeling, which we believe to be one of the most promising yet challenging upcoming fields for food microbiology.
The metabolism of acting microorganisms is largely responsible for the production of safer-to-consume products that are stable, controllable, and have longer shelf lives. The metabolism can be better understood and the fermentation processes can be controlled by GSMMs. For numerous foodrelated microorganisms, such as baker's yeast, lactic acid bacteria (LAB), and most recently acetobacteria, such models have been developed. We can gain insight into the connections between processes by charting the entire metabolism, such as the role of amino acid metabolism in redox balance or the regulation of arginine metabolism in energy metabolism. These models are less used in food biotechnology than they are in chemical biotechnology, where they have become powerful tools for predicting and controlling metabolic engineering and cultivation strategies. Indeed, food contains numerous complications that pose difficulties for metabolic modeling. First of all, foods frequently form solid, opaque, turbid, or multiphase systems.
Quantification of metabolites in complex media, which is necessary to establish precise exchange bounds in GSMMs, is time-consuming, which is an associated obstacle. Alternative readouts or accessible food-matrix models, such as those that have recently been developed for milk, are required to monitor processes in such systems in order to generate data. Lactate (also known as ethanol) is one important product that is used for food preservation. Ironically, the primary modeling technique for GSMMs, traditional Flux Balance Analysis (FBA), has trouble predicting it. Since GSMMs only include stoichiometry and not enzyme kinetics, they require uptake constraints to limit fluxes through the metabolic network. FBA is a problem of optimization: FBA achieves an objective, typically the maximum production of biomass components, by maximizing it. Therefore, high-yield strategies are predicted by FBA by predicting the metabolic activities that produce the most biomass per limited substrate. Even though inefficient overflow metabolism typically occurs in food under nutrient-rich conditions, the FBA problem will always predict respiration over fermentation or acetate formation without additional constraints.
Under food-related conditions, the predictive capabilities of GSMMs have been enhanced by recent modeling advancements. It was demonstrated that lactate formation incurs the least amount of protein expenditure per ATP produced by including resource allocation, or imposing additional constraints to account for the costs of implementing a metabolic pathway. As recently demonstrated for Lactococcus lactis and Saccharomyces cerevisiae, if glucose is abundant, the relevant fitness currency is protein cost rather than metabolic yield. However, balanced growth in a constant environment is required for optimal resource allocation. Mechanistically explaining observed stress responses in Escherichia coli, such as the deactivation of branchedchain amino acid biosynthesis under auxotrophy in oxidative stress, have recently been identified by researchers as metabolic routes that become active in oxidative stress and acid stress conditions. A whole-cell resource allocation model of E. coli was used to incorporate multi-omics (transcriptomics and quantitative proteomics) data.
Food safety might also be made safer by new insights into microbial metabolism. Metabolic models can advance our understanding of the metabolism of other pathogenic or undesirable microbes by following in the footsteps of well-studied classical food-borne human pathogens (like Listeria monocytogenes). A reconstruction of Lactobacillus vini revealed nutrients assimilated into the biomass of this prominent contaminant in sugarcane fermentation, and GSMMs were used to design minimal chemically defined media to cultivate two pathogenic bacteria, Bordetella pertussis and Campylobacter jejuni [1-5].
The biotransformation of indigestible and undisturbed resources into food products is another food science application of fermentation. Coffee and cacao beans are fermented to remove the mucilage layer in traditional biotransformations. Recently, a number of models for members of the cocoafermenting consortium have been developed. A metabolic model of Acetobacter pasteurianus showed how the bacteria first gets its carbon from ethanol and then eats off-flavors like acetoin and acetate. As a means of locating speciesspecific reactions, additional metabolic models of yeast and bacteria were developed. Additionally, this knowledge can be combined with microbial dynamics and metabolite kinetics to create optimal fermentation models, such as those for the fermentation of cacao or coffee.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
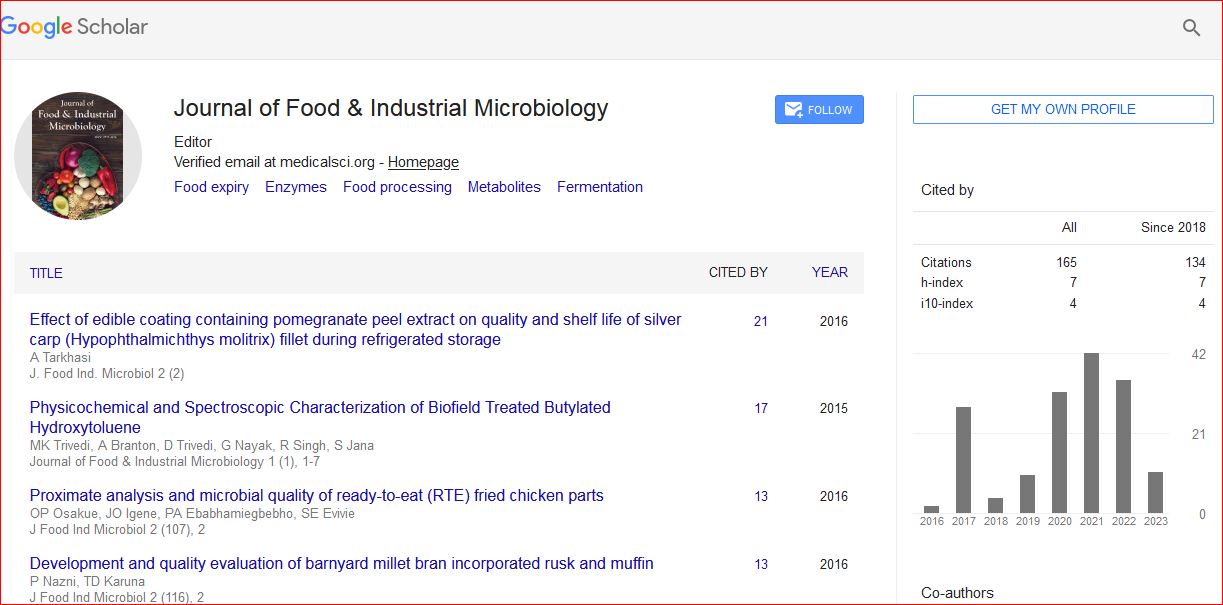
Journal of Food & Industrial Microbiology received 160 citations as per Google Scholar report