Review Article - (2023) Volume 7, Issue 3
Received: 09-May-2023, Manuscript No. jid-23-98310;
Editor assigned: 11-May-2023, Pre QC No. P-98310;
Reviewed: 23-May-2023, QC No. Q-98310;
Revised: 29-May-2023, Manuscript No. R-98310;
Published:
05-Jun-2023
, DOI: 10.37421/2684-4559.2023.7.206
Citation: Carneiro, Sofia, João Paulo Gomes and Rita Macedo. “Nontuberculous Mycobacteria: A Review.” Clin Infect Dis 7 (2023): 206.
Copyright: © 2023 Carneiro S, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Nontuberculous mycobacteria are increasingly causing disease in humans, ranging from skin lesions to widespread disease. Its ubiquitous character in nature makes its exposure very common. For these reasons, diagnosis of the disease, the correct identification/ characterization of the Nontuberculous mycobacteria responsible for the infection, and consequently the definition of the appropriate treatment regimen, remain the major challenge. Treatment is complex, requiring the prolonged use of multiple drugs, which makes it expensive and often brings side effects for the patient. So far, it has not been possible to establish, with certainty, a relationship between in vitro assays and microbiological response to drug treatment, thus making the treatments empirical. Diagnostic and clinical criteria should be updated to enable a more reliable identification in order to improve our understanding of Nontuberculous mycobacteria epidemiology, particularly for the species that have the most potential to cause disease. As an ultimate unavoidable downstream procedure, the use of whole genome data will strongly contribute to Nontuberculous mycobacteria characterization, not only for more precise strain/species differentiation but also eventually to anticipate antibiotic resistance through the identification of resistance markers. With this review, we hope to give the viewer an overview of the Nontuberculous mycobacteria-related topics that we believe are the most important.
Nontuberculous mycobacteria • Epidemiology • NTM disease • Antibiotic resistance
The genus Mycobacterium
First discovered in 1874 by Armauer Hansen, the genus Mycobacterium, with more than 220 species known to date, is the only genus of the Mycobacteriaceae family [1-3]. However, it was just in 1896 that the name Mycobacterium was established by Lehmann and Neumann [4,5].
Bacteria from the genus Mycobacterium are commonly called mycobacteria and, based on their differences in the capacity to grow in vitro, epidemiology, and association with diseases; they can be divided into four different groups: Mycobacterium tuberculosis, Mycobacterium leprae, Mycobacterium ulcerans, and Nontuberculous mycobacteria (NTM) [6].
Mycobacteria are mostly aerobic, non-spore-forming, non-motile, and resistant to acid-alcohol decolorization and colony morphology can vary from rough to smooth and from pigmented to nonpigmented [2,4,7]. One of their most important and unique characteristic is the composition of their cell wall, which is rich in complex lipids and contains long carbon chains (60-90 C) [8-13]. Considering the growth rate, mycobacteria can be classified into two groups: 1) the slow growers (SGM) (require more than one week to be detected on solid media); and 2) the rapid growers (RGM) (require 3-7 days to be detected on solid media). Mycobacteria also have a higher G+C content and lower copy numbers of the ribosomal operon (two copies in the RGM and one copy in the SGM) than most bacteria [14,15].
According to pathogenicity, the genus Mycobacterium can also be subdivided into strict pathogens, opportunistic pathogens, and saprophyte species [16]. Since most of them are opportunistic environmental pathogens, such as saprophytes in soil and water [4], the correct identification in a clinical setting is essential for diagnosis, eventual outbreak detection, and management of the putative underlying disease.
Taxonomy of Mycobacterium genus
Members of the genus Mycobacterium have many phenotypic and genomicbased characteristics that separate them from other genera [17]. In 1957, Ernest Runyon proposed the first taxonomic division of mycobacteria into four groups based on the growth rate and production of a pigment [18,19]. According to this scheme, SGM are divided in three groups: group I or photochromogen (pigmented when exposed to light), group II or scotochromogen (always pigmented) and group III or nonphotochromogen (nonpigmented). The RGM belong to group IV [7,18,20]. Of note, members of the M. avium complex are considered nonphotochromogen, however, some isolates are capable to produce slightly pigmented colonies [7] (Table 1).
| Runyon Group | Classification according to Growth Rate |
Classification according to Pigmentation |
Description | Organisms (e.g) |
|---|---|---|---|---|
| I | Slow grower | Photochromogen | Cultures non pigmented in the dark and pigmented when exposed to the light |
M. kansasii M. marinun |
| II | Slow grower | Scotochromogen | Pigmented colonies in either dark or light incubated cultures |
M. gordonae M. scrofulaceum |
| III | Slow grower | Nonphotochromogen | Nonpigmented colonies either in cultures incubated in the dark or exposed to light |
M. avium complex M. tuberculosis |
| IV | Rapid grower | - | Pigmented or nonpigmented | M. abscessus - chelonae complex M. fortuitum |
Despite some limitations, the analysis of the 16S rRNA encoding gene, which is mostly conserved between species, was used to better differentiate some Mycobacterium species [14,21] and supported Runyon's classification into RGM and SGM, which still continues to be used by mycobacteriologists [6,20]. However, the need to improve the robustness and discrimination between different species in phylogenetic trees led to the analysis of concatenated sequences of housekeeping genes such as the 65-KDa heat shock protein gene (hsp65), RNA polymerase β- subunit (rpoB) and DNA gyrase subunit B (gyrB) [22-24]. No major breakthrough was accomplished with these approaches and the introduction of whole genome sequencing (WGS) coupled with bioinformatics tools revolutionized mycobacteria classification. Although genome-based phylogenies often do not produce very different tree topologies compared to conventional (phenotype-based) reconstructions, it is now clear that phylogenetic studies that ignore all genome-based analyses are outdated as they lack discriminatory power [25]. The first two studies relying on WGS data improved our understanding of mycobacteria taxonomy by validating the evident distinction between RGM and SGM: RGM assume the most ancestral branch, being the members of Mycobacterium abscessus-chelonae complex the most ancestral cluster, whereas species belonging to M. terrae complex occupy an intermediate position and SGM are clearly separated in another branch [17,25] (Figure 1).

Figure 1. Phylogenetic tree of Mycobacterium species.
Given the rampant advances in genomics and with the permanent release of new WGS data, it is now believed that the taxonomy of mycobacteria is well established. Still, one must realize that the genus phylogenetic tree is dynamic due to the continuous application of more robust differentiation and identification methods, leading, for example, to the inclusion of new species. In this regard, new studies have assumed some controversial opinions about the definitions of complexes/clades within the genus Mycobacterium. Gupta RS, et al. [15] defended the redistribution of the members of the genus Mycobacterium into five new groups: “Tuberculosis-Simiae clade” that includes all of the major human pathogens, and four novel genera: the SGM Mycolicibacterium gen. nov. (Fortuitum-Vaccae clade) and Mycolicibacillus gen. nov. (Triviale clade), and the RGM Mycobacteroides gen. nov. (Abscessus-Chelonae clade) and Mycolicibacter gen. nov. (Terrae clade) [15].
In contrast, Tortoli E, et al. [26] opts for the classical nomenclature that includes 192 species, of which five (M. chelonae-abscessus complex, M. fortuitum, M. avium, M. leprae and M. tuberculosis) are divided into subspecies [26,27].
Since this is still the most widely used approach in the scientific community, it will also be used in this review.
Nontuberculous Mycobacteria
Nontuberculous mycobacteria, or just NTM, are also known as mycobacteria other than tuberculosis (MOTT), atypical mycobacteria or environmental mycobacteria [6,11]. These bacteria often exhibit saprophytic, commensal, and symbiotic behaviors, and they are classified as opportunistic human pathogens [28,29]. They have the capacity to tolerate a wide temperature range, do not grow on standard culture media, and are resistant to many antibiotics and disinfectants [30]. The cell wall, which is rich in lipids making it hydrophobic and impermeable, along with the slow growth rate, are the major characteristics that allow mycobacteria to persist in extreme environments [28,31,32]. As such, NTM have several reservoirs, either natural or human-made, such as soils, water, dust and air [32-41]. In addition, they also have the ability to infect animals, which gives them importance in both human and veterinary medicine [39,42,43].
Unlike M. tuberculosis, the transmission of NTM does not seem to be personto- person but rather trough inhalation of aerosols from infected sources and, in some cases, ingestion or trauma events [44]. Although it has been suggested that person-to-person transmission may occur between patients with cystic fibrosis [45,46], this seems to be an exception [47].
The environment is rich in RGM and most of them are not associated with disease. However, some species such as Mycobacterium abscessus-chelonae complex (MABC) (M. abscessus spp abscessus, M. abscessus spp. bolletii and M. abscessus spp. massiliense) and Mycobacterium fortuitum can cause disease Among the SGM, the ones that are most commonly isolated and associated with disease are: Mycobacterium avium complex (MAC), particularly the sub-species M. avium spp. avium, M. avium spp. intracellulare and M. avium spp chimaera, Mycobacterium xenopi and Mycobacterium kansasii [30,48-50].
As NTM are ubiquitous in environment, exposure is frequent [51,52], so the detection of these opportunistic pathogens in a clinical sample is not enough to classify them as the disease causing agents, making it difficult to clarify the clinical significance of NTM [53].
Epidemiology of Nontuberculous mycobacteria
Although many studies report an increase in NTM human disease, and these are already recognized as a global health concern [54-57], the epidemiology of NTM is not well understood [58,59]. This lack of knowledge can be attributed to several factors: 1) NTM disease is not of mandatory reporting, so most of the cases are not reported to public health authorities and, as a consequence, incidence data relies just on the number of laboratory isolates [11,36,60]; 2) diagnosis of NTM requires considerable time and may be misdiagnosed as tuberculosis or other lung disorder [61-63]; and 3) the existing diagnostic tests are unreliable and costly and no prognostic tests are available [30].
The distribution of NTM species seems to be different among geographic regions and populations [36]. Hoefsloot et al. [49] showed that MAC is predominant worldwide, M. avium is predominant in North and South America and Europe, and M. intracellulare is most frequently isolated in Africa and Australia. They also described that M. xenopi is particularly prevalent in Hungary, Croatia, Northern Italy, Ontario, and in the area close to the English Channel. M. kansasii appears to be more prevalent in South America, Eastern Europe and the metropolitan centers of London, Paris and Tokyo and some areas of South Africa. M. malmoense seems to predominate in Europe [49].
More recently, Farnia P, et al. [64] published a systematic review on the distribution of NTM [64]. In the north and east of Europe, the most frequently found NTM in clinical samples belonged to MAC (in particular M. avium) and M. gordonae. In the south and west of Europe, the most common NTM isolated were M. xenopi, M. gordonae, and MAC, mainly M. avium. Similarly to the study of Hoefsloot et al. [49], Farnia P, et al. [64] also observed that in Africa and Australia M. intracellulare was the most frequent NTM isolated, followed by M. avium, and M. kansaii in Africa and by M. fortuitum in Australia. Furthermore, according to this study, in East Asia the most frequent NTM species isolated are M. avium and M. abscessus, in west Asia, M. fortuitum, M. gordonae, and M. simiae and, in south Asia, M. fortuitum, M. chelonae and M. xenopi [64].
A recent study from Portugal showed that MAC is the most isolated NTM associated with disease, followed by MABC and M. fortuitum. As expected, the majority of the cases appeared in the regions with higher population density [60,65].
Of note, a common denominator of all of these epidemiological studies is that the prevalence of the disease likely caused by NTM seems to increase with the aging of the population, with the development of laboratory techniques, with the increase in the number of immunosuppressed patients, and with a decrease in the incidence of tuberculosis [49,50,66-68]. Thus, it is clear that there is a need to create a broad approach involving both the collection of informative clinical data and more robust laboratory procedures in order to be able to estimate the true impact of these infections [1,69,70].
Pathogenesis and immune response in NTM infections
Although exposure to these bacteria is frequent, NTM disease is relatively uncommon, leading to the assumption that normal host defense mechanisms are sufficient to prevent infection. This also suggests that patients who develop NTM disease must have specific susceptibility factors that make them vulnerable [52]. However, the physiologic and cellular conditions that likely facilitate NTM infections are poorly understood [71]. On the pathogen side, with the exception of mycolactone of M. ulcerans, there is no evidence of specific virulence factors among mycobacteria that may potentiate the infection [11]. It has recently been suggested that the capacity of some NTM (e.g: MAC, Mycobacterium abscessus and Mycobacterium kansasii) to change their colony morphotype from smooth to rough, contributes to their virulence, being the latest the most virulent in most of the cases, with exception of MAC where smooth colonies are the most virulent [11,72-74].
M. kansasii affects mostly the lungs but it can also cause infections in lymph nodes, bone, skin and, in cases where the host has low CD4 counts (below 50/ mm3), disseminated disease can be developed [75-77]. This NTM species has the ability to enter macrophages, which in turn are the ideal environment for the microorganism to develop and be transported to other tissues[77,78].
M. abscessus is the most virulent fast-growing mycobacteria [79]. The infection shares similarities with the one caused by M. avium and M. tuberculosis (i.e., formation of granulomas and persistence of infection) however, its mechanisms of transmission and establishment of disease are still not well understood [79,80]. It is known that the colony morphotype plays an important role in the ability of this NTM species to infect, as it allows the transition between a phenotype with a greater ability to colonize (smooth) and a more virulent and invasive phenotype (rough) [81]. Both smooth and rough phenotypes can survive inside the macrophages and be maintained in loner phagosomes. The difference between these two phenotypes is that the smooth variant is usually able to prevent the activation of apoptosis and autophagy. On the other hand, when the transition to the rough variant occurs, bacterial cords formation and acidification of the phagosome begin, resulting in a massive tissue destruction leading to severe infection. The production of bacterial cords only happens in rough forms and is determinant to establish the infection [80],[82-84].
Concerning MAC infections, rough morphotypes are less pathogenic than the smooth ones, as they lack genes for glycopeptidolipids synthesis that are significant determinants for virulence [85-87]. These are necessary to biofilm production, which is an important mechanism to disrupt the host's immunity leading to settlement facilitation and resulting in invasion of bronchial epithelium [77,88]. MAC species also have the ability to survive the acidic pH of the stomach, overcome the acid barrier and gain access to the intestinal lumen [77]. Nevertheless, some differences can be found within the species of MAC.
The phagocytes, after engulfing mycobacteria, activate a series of complex cascade reactions [89]; cytokines, such as interleukin-12 (IL-12), interferon-gamma (IFN-γ) and tumor necrosis factor-α (TNF-α), play a role in antimycobacterial immune response and regulation [90]. IL-12 production leads to natural killer (NK) cells production and t-lymphocyte proliferation [91], which are essential for the innate immune response to M. avium infections [92]. In fact, the secretion of TNF-α, IFN-γ and granulocyte-macrophage colony-stimulating factor (GM-CSF), will stimulate infected macrophages enabling them to control the intracellular infection [77,93,94]. Regarding M. abscessus infection, the rough morphotype prevents macrophages innate response [79,95,96]. In these cases, the immune innate response is activated by the interaction with toll-like receptor 2 (TLR2) resulting in a TNF-α release. IL-12 is also released leading to an activation and polarization of naive CD4+ T cells towards Th1 cells to produce IFN-γ [79]. The adaptive immune response also plays an important role in the immune response against M. abscessus infections, as it is important for granuloma formation through recruitment of B and T cells [97]. The granuloma is a structure that is formed during mycobacterial infections and usually contains the infection. However, as already mentioned, M. abscessus has the ability to change from smooth to rough colonies resulting in granuloma collapse and dissemination of the infection [84].
Manifestation of the disease is the reflex of an interaction between the exposure (for example, the infecting dose and duration of exposure), the microorganism (pathogenicity and virulence), and the host (immune status, genetic risk factors and prior lung disease) [62,98,99]. The respiratory tract is the most frequent target of NTM, however, these bacteria have the ability to infect a wide variety of body sites [100]. The most common diseases caused by NTM are pulmonary disease, lymphadenitis, skin, soft tissue, and bone disease and disseminated disease in severely immunocompromised patients (Figure 2) [53,101,102].

Figure 2. Clinical diseases more common caused by NTM.
A correct and early identification of NTM is essential for the management of the disease [11]. Since the symptoms of NTM are non-specific, there is often a delay in diagnosis, resulting in disease progression and, eventually, more complicated treatment regimens with more side effects [11,103].
Pulmonary and extrapulmonary NTM diseases are managed separately, although clinical, radiological and laboratory findings are critical in both cases [104]. In either case, the diagnosis is complicated as NTM exposure is very common, although disease development is rare [105]. Furthermore, due to the ubiquitous character of NTM, in most cases of pulmonary disease one must take into account that the presence of a NTM positive culture from respiratory sites only reflects colonization rather than the detection of the disease-causing agent. In addition, the respiratory samples are non-sterile which makes it another factor that creates uncertainty in the final diagnosis [104].
Although NTM lung disease does not reveal specific symptoms (i.e., cough, fever, fatigue, weight loss and hemoptysis), the radiological findings are suggestive of this type of infection. Typically, specific bronchiectasis with nodules or cavitation is found in nodular bronchiectasis disease and fibrocavitary disease, respectively [54,106,107].
Lymphadenitis, with cervical adenitis as the clinical presentation, is common among immunocompetent children [11,53,104,108]. The first symptom that is suggestive of lymphadenitis is unilateral indolent swelling that persists in time without systemic symptoms associated [104,108]. The definitive diagnosis relies on the microbiological findings through lymph node sample analysis as these also allow excluding tuberculosis and pyogenic lymphadenitis [104,108].
Skin, bone, and soft tissues disease, as the other forms of NTM infections, also demand clinical context, microbiological results, histopathology results compatible with the diagnosis and history of underling disease [53,104]. Skin and soft tissues disease may develop in immunocompromised patients after a puncture wound or a traumatic injury [108,109]. Nosocomial infections are acquired due to invasive therapeutic interventions, surgeries or long term intravenous catheters [108]. Albeit rare, NTM have been isolated from osteomyelitis cases and, in immunocompromised patients, is often a result of disseminated disease [110]. Bone infection can occur in immunocompetent patients after an accidental trauma or surgery [111,112]. Diagnosis is difficult manly due to the lack of specific examination methods and to the various possible clinical presentations [111]. Magnetic resonance is used to help find possible sites of infection; however, it cannot clearly differentiate infection caused by NTM or TB [113]. As such, for a correct diagnosis, microbiological and/or histological confirmation is mandatory [104].
Disseminated disease may appear in patients with low CD4+ T cell counts [53] and, once more, symptoms are nonspecific (e.g., fever, weight loss and abdominal pain) [104]. A positive blood culture is the main differential diagnosis, but it can also be done by the isolation of NTM from another sterile sample, for example lymph node, bone marrow or liver [104].
Some genetic/ heritable and acquired disorders that compromise the lung may act as risk factors to NTM disease. Examples of acquired disorders are smoking-related emphysema, bronchiectasis as a result of another unrelated infection, use of immunosuppressives, pneumoconiosis and chronic aspiration [114-116]. On the other hand, genetic disorders include cystic fibrosis (CF), elastin deficiency, congenital bronchial cartilage deficiency, alpha-1-antitypsin deficiency and, primary ciliary dyskinesia [117-121]. Several studies indicate that NTM infections are more common in female gender, and consider older age and low body weight as risk factors for disease [122-126]. In addition, an immunosuppressed status, as the one associated with human immunodeficiency virus infection, transplantation, or defects in the pathways of the IL-12, TNF-α and, IFN-γ are well-known risk factors [71,108] [127-130]. Also, in an infected individual, an immunosuppressive treatment increases the risk of progression to disease by NTM.
The clinical significance of NTM isolated in the airways is hard to measure since they are environmental bacteria, and colonization of the airways is common. For this reason, the American Thoracic Society (ATS) and the Infectious Disease Society of America (IDSA) have provided criteria for NTM disease diagnosis. It requires microbiological, clinical, and radiographic correlation, with all of these criteria of equal importance [53]. As a NTM-positive respiratory specimen from a patient who has no evidence of underlying disease may imply contamination, at least two separate positive cultures of sputum specimens collected in different time periods are required to define the case as possible disease. However, if a patient has a lung biopsy with mycobacterial histopathological features and a positive NTM culture is isolated from the biopsy sample, a single positive result is enough to consider as a potential case of disease. The same criteria apply if the patient has a positive culture of other invasively collected biological samples such as bronchial or bronchoalveolar lavage. Clinical criteria for case definition include pulmonary or systemic symptoms such as cough, sputum production, chest pain, fatigue, fever, and weight loss whereas radiographic criteria are based on chest radiographic results showing nodular or cavitary opacities or computerized tomography (CT) scan results showing bronchiectasis with multiple nodules. Even if a patient meets all the criteria necessary to be classified as having NTM disease, the pathogenicity of the NTM isolate must be taken into account, and for low-pathogenic NTM species, recurrent positive cultures and strong clinical and radiological evidence are required for establishment of NTM disease [53,61,131].
Treatment regimens used in NTM infections require the prolonged use of multiple drugs, which is costly and may cause severe side effects for the patient, as these drugs usually show high toxicity [61]. As such, each treatment is unique and dependent on the individual comorbidities of each patient and the risk-benefit ratio [132]. The decision of which combination of drugs to use depends on the species and, in some cases, the subspecies of the NTM causing the disease. Although drug susceptibility testing (DST) is advised for correct treatment regimen design, correlation between in vitro and in vivo outcome is not yet well established [133,134] and, as such, treatments are still mostly empirical.
Treatment regimens for SGM usually include ethambutol, rifampicin and a macrolide and, in some severe cases, amikacin or streptomycin can also be added. Not every SGM species has defined breakpoints for DST. Members of MAC are tested for clarithromycin, linezolid, and moxifloxacin; M. kansasii is tested for clarithromycin, rifampicin, amikacin, ciprofloxacin, ethambutol, isoniazid, linezolid, moxifloxacin, rifabutin, streptomycin, and trimethoprimsulfamethoxazole; and M. marinum is tested for amikacin, ciprofloxacin, clarithromycin, doxycycline, ethambutol, moxifloxacin, rifabutin, rifampin, and trimethoprim-sulfamethoxazole [135].
In case of infections with RGM, the treatment regimen is based on DST results that are performed for macrolides, fluroquinolones, amikacin, imipenem, tetracyclines, linezolid, and trimethoprim-sulfamethoxazole [53].
Mechanisms of antimicrobial resistance
Antimicrobial resistance is a global problem underlying infections caused by bacteria, and mycobacteria are no exception. Most NTM species are resistant to a wide array of antibiotics and both natural and acquired resistances are important determinants for the treatment success [53,136]. DST is used to determine the interplay between resistance and susceptibility during suboptimal drug exposure and selection, which helps in the establishment of appropriate treatment regimens [136]. As NTM have natural/intrinsic resistance according to species or subspecies, it is of extreme importance to identify the NTM causing the disease in order to define the appropriate treatment regimen, even if an empiric one [53]. This natural drug resistance is conferred by mechanisms associated with the cell wall permeability, thickness, formation of granulomas or the capacity to form biofilms. These mechanisms interfere with drug uptake that allow their biotransformation, or decrease the affinity with the drug target [136-141]. The highly hydrophobic and impermeable cell wall of mycobacterial cells, which is composed by N-glycolyl muramic acid and is abundant in lipids constituted by long chain fatty acids with more than 90 carbons [139-141], hampers the diffusion of hydrophilic antibiotics and nutrients through this layer [136,142]. For these reason, the transport mechanisms across membrane is essentially controlled by porin channels [136,142,143]. It is the activity of these porins that determines the susceptibility to hydrophilic (e.g norfloxacin and β-lactam) and hydrophobic (e.g vancomycin and rifampicin) antibiotics [144-146]. In addition, NTM also possess efflux systems (e.g. P55, tetV and tap that confer resistance to aminoglycosides and tetracyclines) that avoid the accumulation of drugs inside the cell [136],[147- 149]. These efflux pumps are substrate-specific, but others transport a wide range of substrates, conferring resistance to multiple drugs at once [150]. Another efflux pump system has been described in MAC: MmpL5/MmpS5, which confers resistance to clofazimine and bedaquiline.
The persistence of a multidrug-resistant phenotype is possible due to the existence of several genes and systems involved in cell wall maintenance such as the protein kinase G (pknG), and asnB in M. smegmatis, and the mtrAB in M. smegmatis and M. avium [143]. Whenever a disruption in these genes is observed, there will usually occur a decrease in hydrophobicity of the mycobacterial cell wall, leading to increased susceptibility to lipophilic antibiotics (e.g. macrolides, rifamycins, and penicillins) [151-155].
As mentioned before, the formation of biofilms and granulomas are two additional mechanisms to promote antimicrobial resistance in NTM [138,156]. The lipid-rich extracellular matrix of the biofilm works as a barrier, which does not allow the penetration of drugs [156]. Furthermore, in biofilms, horizontal gene transfer is potentiated due to the interactions between the bacteria that form the biofilm layers, promoting the spread of drug resistance [157]. Moreover, some genes are expressed differently when the bacteria grows in biofilms [158]. For example, it is assumed that the increased chlorine resistance noted in M. avium and M. intracellulare cells when these bacteria grow in biofilms is due to changes in the cell wall, which in turn results from changes in the structure of mycolic acid [159].
Changes in gene expression may lead to modifications in the binding sites of antibiotics. This is the case of the erythromycin ribosomal methylase (erm) gene that confers resistance to macrolides by methylating the bacterial ribosome, thereby blocking the binding site for macrolides [160,161]. The erm gene has been described in M. abscessus spp. abscessus, M. abscessus spp. bolletii, M. fortuitum, and M. porcinum [133,162,163]. In M. abscessus spp. massiliense the treatment with macrolides can still be an option as inducible macrolide resistance does not occur in this subspecies [164,165].
On the other hand, acquired resistance appears mainly due to spontaneous mutations in chromosomal genes, especially during antibiotic treatment [137,150] and, due to this type of resistance, multidrug-resistance pathogens are rapidly increasing worldwide [166]. Mutations in 23S rRNA gene (rrl) and in 16S rRNA gene (rrs) are responsible for resistance to macrolide and aminoglycoside respectively, in MABC and MAC clinical isolates [156,162][167-170]. Rifamycin resistance is acquired by a mutation in the rpoB gene encoding the β-subunit of RNA polymerase resulting in a blockage of RNA synthesis [171,172]. These mutations have been reported in MAC and M. kansasii [133,173,174]. The expression of arr gene in M. smegmatis and M. abscessus it is also associated with a reduced efficacy of rifamycins such as rifampicin and rifabutin [175,176].
Infections caused by nontuberculous mycobacteria are becoming a public health problem. Although they are more common in immunosuppressed patients and with other comorbidities, there are also reports of immunocompetent patients infected with NTM disease. In order to increase the knowledge on pathogenicity, evolution, treatment, resistance mechanisms, and environmental niches, studies on NTM are mandatory. This study summarizes the NTM-related issues that we consider the most important.
Sofia Carneiro: Literature review; writing draft preparation. Rita Macedo and João Paulo Gomes: Reviewing and editing. All authors have read and approved the final version of the manuscript.
None.
None.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
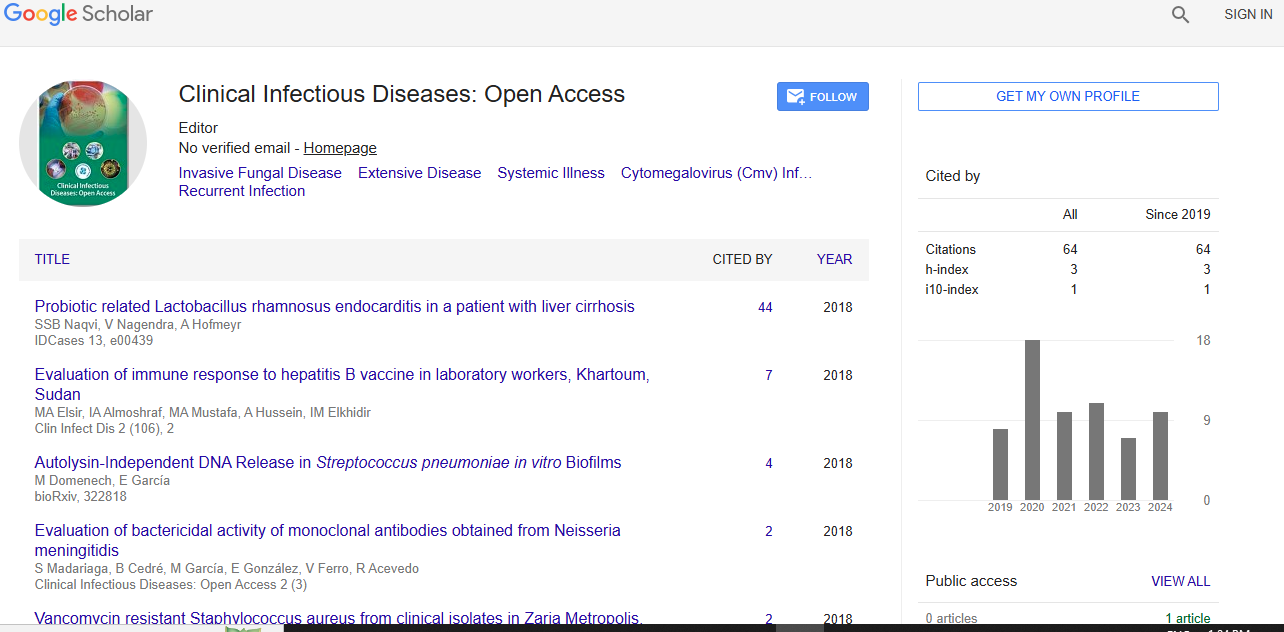
Clinical Infectious Diseases: Open Access received 1149 citations as per Google Scholar report