Mini Review - (2021) Volume 12, Issue 9
Received: 18-Sep-2021
Published:
08-Oct-2021
, DOI: 10.37421/2157-7099.21.12.596
Citation: L Aravind . "Origin and Evolution of Eukaryotes." J Cytol Histol 12 (2021): 596.
Copyright: © 2021 L Aravind. This is an open-access article distributed under the
terms of the Creative Commons Attribution License, which permits unrestricted
use, distribution, and reproduction in any medium, provided the original author
and source are credited.
The endosymbiotic beginning of eukaryotes united two dissimilar genomes in the cell. Also, eukaryotic normal history has included other endosymbiotic occasions, phagotrophic utilization of living beings, and personal associations with infections and endoparasites. These wonders worked with enormous scope sidelong quality exchange and organic contentions. We combine data from almost twenty years of genomics to outline how the exchange between horizontal quality exchange and natural contentions has affected the rise of new variations in eukaryotes. Utilizing apicomplexans as model, we outline how sidelong exchange from creatures has added to extraordinary parasite-have interfaces included grip and O-connected glycosylation-related spaces. Variations, arising because of extraordinary choice for variety in the atomic members in organismal and genomic clashes, being scattered by sidelong exchange, were accordingly exapted for eukaryote-explicit developments. We show this utilizing models identifying with eukaryotic chromatin, RNAi and RNA-preparing frameworks, flagging pathways, apoptosis and invulnerability. We feature the significant commitments from synergist areas of bacterial poison frameworks to the beginning of flagging catalysts (e.g., ADP-ribosylation and little particle courier blend), mutagenic chemicals for invulnerable receptor broadening and RNA-preparing. Also, we examine commitments of bacterial anti-infection or siderophore combination frameworks and intra-genomic and intra-cell childish components (e.g., limitation change, portable components and lysogenic phages) in the rise of chromatin renovating/adjusting chemicals and RNA-based guideline. We foster the idea that organic struggle frameworks filled in as transformative "nurseries" for advancements in the protein world, which were conveyed to eukaryotes through parallel quality stream to spike key developmental advancements right from nucleogenesis to genealogy explicit variations.
Antibiotics • Biological Conflict • Endosymbiosis • Immunity Pr oteins • Restriction-Modfication • Toxins
Since the time the rise of the endosymbiotic theory as the essential model for the beginning of eukaryotes there has been considerable interest in two significant issues which it delivered, in particular enormous scope sidelong quality stream and hereditary struggles. While the specific subtleties of the idea of this endosymbiotic occasion are as yet discussed, by its very nature the endosymbiotic theory infers quality stream between the alpha proteobacterial mitochondrial progeni-pinnacle and the nucleo-cytoplasmic begetter of archaeal parentage [1-3]. This wonder isn't only pertinent to the beginning of eukaryotes, yet in addition a few other symbiogenic occasions that molded the resulting advancement of eukaryotes, like the beginning of the essential photosynthetic eukaryotes, including the plants, and the various optional or tertiary photosynthetic eukaryotes [4,5]. In the previous occasion, not simply the notable quality stream from cyanobacteria, yet additionally corresponding commitments from a chlamydia-like endosymbiont have been hypothesized [6]. Furthermore, there are other between organismal connections that have happened all through eukaryotic evolution, which have brought about similar quality stream, though in a more rambling style. Eukaryotes are described by a wide-scope of close organismal associations. For sure, cytoplasmic advantageous microbes, practically identical to the ancestors of the mitochondria and chloroplasts, and contamination by a few kinds of enormous DNA infections are a typical element of numerous eukaryotes, including delegates of the metazoan and amoebozoan heredities [7]. There are likewise instances of a few Maybe emotional between eukaryotic affiliations, similar to endoparasitism as shown by apicomplexans, karyoklepty, or "burglary" of chlorophyte cores (alongside the chloroplasts) saw among ciliates, or karyoparasitism, including infusion of parasitic cores into have cells, which is seen in certain rhodophytes [8]. Further, it has been noticed that the phagotrophic nutrition of numerous eukaryotes can likewise bring about a more broad type of hereditary chimerism, worked with by the steady engulfment of hereditary material of specific kinds of microorganisms and eukaryotes. However different eukaryotes, like the rotifers, seem to even effectively take part in take-up and joining of hereditary material from their surroundings notwithstanding the proposed job in making up for the absence of sexual proliferation, this phenomenon likewise fills in as a course for outstanding "outsider" quality stream. In this way, it has become progressively clear in the beyond twenty years that quality stream between far off ancestries and the ensuing genomic chimerism may play an eminent part in the development of eukaryotes.
Between organismal and intra-organismal hereditary struggles are ordinary elements across all authoritative degrees of life (9). In their easiest structure they incorporate different trophic connections between creatures, like predation. Such contentions may likewise emerge between various cells of similar species agreeably aggregating to frame a multicellular gathering or creating as a multicellular creature due the rise of "miscreants," whose hereditary interests don't line up with the leftover participating cells. At the level of a solitary cell, as the between ests of various genomes living inside it are not really lined up with one another, there is potential for one more degree of hereditary struggles [9]. Such contentions have a long transformative history in the prokaryotic superkingdoms as the cooperations among plasmids and the cellular genome. Be that as it may, the endosymbiotic beginning of eukaryotes made it one of their quintessential provisions since it united various particular genomes (i.e., the atomic and mitochondrial) in a solitary cell. Such between genomic clashes inside the cell hide their extended in course of eukaryotic advancement due to additional affiliations presenting connections with genomes from plastids, nucleomorphs, and endosymbiotic/ parasitic and intra-cell bacterial hunters of mitochondria. In a few cases advantageous microorganisms are engaged with staggered participation struggle connections: For example, the bacterial symbiont Photorhabdus empowers ruthless nematodes to benefit from creepy crawlies by killing them with poisons, while the endosymbiotic bacterium Hamiltonella defensa secures aphids against parasitoid wasps by conveying toxins against them. Clashes between the phone genomes and infections that exploit them for their own proliferation add one more measurement to clashes happening inside cells. At long last, there may be hereditary contentions inside a solitary genome itself, emerging from a wide assortment of narrow minded components attempting to maximize their own wellness to the detriment of the leftover qualities [2]. These self-centered components are frequently portrayed by a level of intra-or potentially between genomic mobility and accept a befuddling exhibit of structures, including various particular kinds of transposable components, limitation alteration, and poison serum frameworks [2,9]. The previous components catalyze or work with their own multiplication, while the last components authorize cellular genomes to hold them by killing cells in which they are disturbed. In spite of being fundamentally childish components, they may on events give a wellness benefit to genomes, as this indirectly increases their own wellness [9].
These struggles are frequently straightforwardly intervened by specific particles, either proteins or little atoms which go about as "chemical deadly implements"; albeit in multicellular structures it very well may be reflected as morphological elements that fill in as weaponry [2,4]. As anyone might expect, every one of the many degrees of organismal struggle have started off serious "arms races" between the cooperating living beings, whose marks are regularly found as broad enhancement of the proteins straightforwardly partaking in, or measuring particles sent in struggle. Associatively, there is additionally a comparative quick diversification of proteins straightforwardly engaged with shielding or filling in as remedys against the substance deadly implements sent in the conflict. Significantly, both the hostile and guarded atomic variations engaged with these contentions can be communicated between genomes via sidelong exchange and is a significant factor both in the spread of anti-infection creation and obstruction among prokaryotes.
The always extending genomic information from the two eukaryotes and prokaryotes, alongside genome-scale investigation, has significantly explained the significant patterns in the genomic chimerism emerging from the bacterial and archaeal forebears of the eukaryotes [1,2]. These investigations have especially separated the cell frameworks which have a fundamentally archaeal provenance (e.g., center DNA replication, center RNA digestion, and interpretation) as against those with a basically bacterial provenance (different parts of energy, anabolic, and catabolic digestion). In any case, revealing the starting points of explicit frameworks, which give off an impression of being eukaryotic synapomorphies (or shared inferred characters), have required a fairly unmistakable computational methodology depending on top to bottom examination of protein groupings and structures [2]. Such investigations uncovered looks at an intrigue between quality move through sidelong exchange and the particular powers following up on molecular players in organismal and intra-genomic struggle in forming the advancement of key parts of frameworks, for example, eukaryotic chromatin, RNA-based quality guideline, and certain signing pathways. Notwithstanding, this part of eukaryotic advancement is impressively overlooked. Thus, in this article we present an engineered outline of:
1) How enormous scope horizontal quality stream between cooperating creatures has worked with the rise of new variations sent in between organismal clash.
2) How transformations created because of the extraordinary determination for variety in the sub-atomic members in organismal and genomic conflicts were scattered by parallel exchange and hence exapted for different eukaryote-explicit variations.
Because of impediments of room, we don't give a complete study of all known occasions of the above measures. All things being equal, we endeavor to high-light the significance of these cycles in the development of key transformations, in early eukaryotes, yet in addition during their ensuing advancement, with assorted outlines rising up out of late examinations. We should stress that in this article we basically utilize distributed models that have been accounted for in several individual examinations on different natural frameworks or protein families. Be that as it may, this is whenever they initially are being united to make a sound picture. An itemized show of the methodological contraption for grouping, structure and phylogenetic examination of the introduced models is blocked because of constraints of room. Nonetheless, we allude perusers to the singular examinations from which we draw our models for insights about the computational investigation of the proteins considered here. We utilize these to foster a calculated system for understanding the significance of the differentiating powers acting during biological clashes in working with transformations that assumed a part in the alleged "significant advances" of eukaryotic developments [10].
The reason why proteins derived from biological conflict systems appear to be recruited for other functions might be attributed to the consequences of natural selection. Not surprisingly, toxin-immunity systems used in inter-organismal conflict have a large effect on the fitness of both the organisms producing toxins and those defending against them, thereby escalating an arms race situation. Many of the conflict systems deployed by bacteria might even function at the interface of symbiotic and parasitic interactions of bacteria and eukaryotes, thereby developing adaptations to effectively target components of eukaryotic systems. Toxins and immunity proteins of intra-genomic selfish elements are also under multiple levels of selection that foster their diversification. At one level they are under selection to evade host resistance to function effectively as addictive agents. At another level many of them might also be under selection to function as effective stress response mechanisms that allow their host genomes to survive adverse conditions. Consequently, there are strong selective pressures for constant diversification of toxins and the corresponding immunity proteins in various conflict systems. Hence, these biological conflicts could have functioned as evolutionary “nurseries” for innovations in both prokaryotic and eukaryotic proteins. Hence, lateral gene flow from symbionts, parasites and other modes of DNA uptake has enabled eukaryotes to have access to and import a “readymade” set of molecular innovations from such biological conflict systems. When recruited in non-conflict biological contexts, they can in turn spur the emergence of new interactions in eukaryotic systems. Thus, number of key eukaryotic innovations can be traced back to the above-described players in biological conflict systems, such as secondary metabolism operons, RM, polymorphic and host-directed toxins systems, anti-phage systems, phage counter- restriction strategies, and mobile elements. These systems appear to have particularly expanded in bacteria on account of the presence of operons, extensive lateral transfer with several modes of DNA uptake and recombination, perhaps combined with high effective population sizes. Thus, organismal and genomic conflicts as the basis for major molecular innovations, which in turn might facilitate major evolutionary transitions, can be considered a general evolutionary principle.
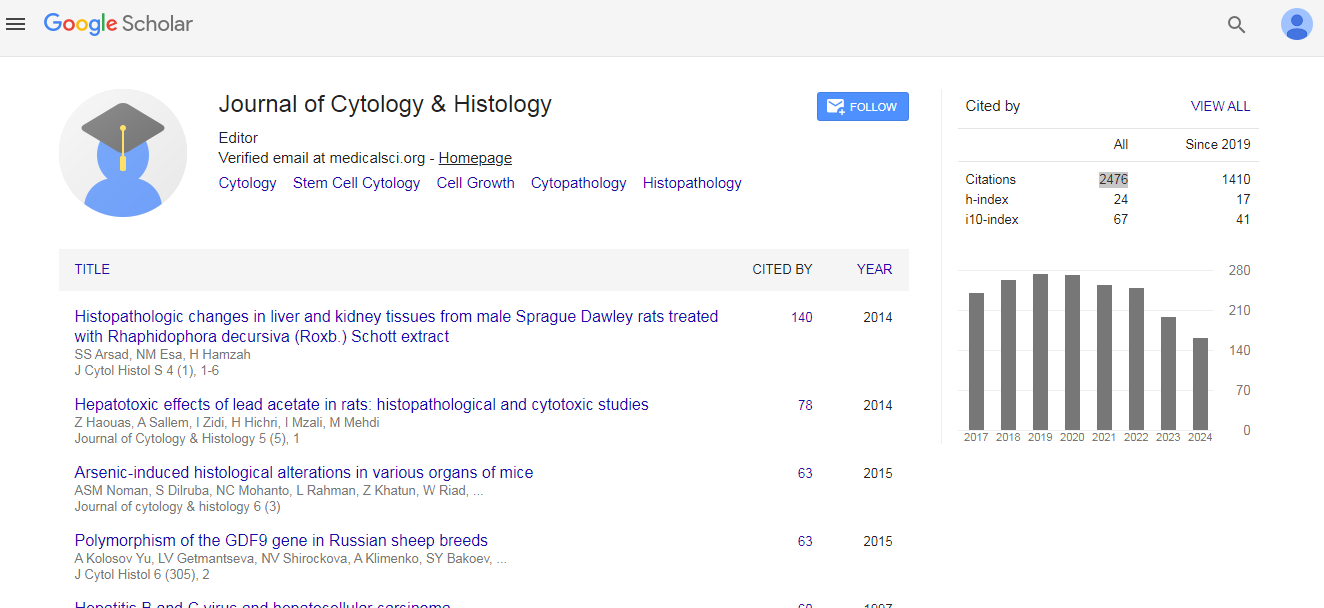
Journal of Cytology & Histology received 2476 citations as per Google Scholar report