Perspective - (2022) Volume 7, Issue 2
Received: 08-Mar-2022, Manuscript No. jib-22-60248;
Editor assigned: 10-Mar-2022, Pre QC No. P-60248;
Reviewed: 24-Mar-2022, QC No. Q-60248 ;
Revised: 29-Mar-2022, Manuscript No. R-60248;
Published:
05-Apr-2022
, DOI: 10.37421/2476-1966.2022.7.170
Citation: Pullela, Sushma. “Pre-erythrocytic Stages of Malaria: Immune Responses.” J Immuno Biol 7 (2022): 170. DOI: 10.37421/2476-1966.2022.7.170
Copyright: © 2022 Pullela S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Sources of funding : None
During a blood meal, Plasmodium spp.-infected mosquitos inject sporozoites into the skin of a mammalian host. These go into the circulatory system of the host and cause an infection in the liver. Merozoites infiltrate the bloodstream after a quiet metamorphosis, resulting in the symptomatic and transmissible stages of malaria. The silent pre-erythrocytic malaria stage is a disease bottleneck where chemotherapeutic and immunoprophylactic therapies can stop the disease from progressing to clinical malaria. RTS,S/ AS01, the only malaria vaccine on the verge of approval, despite its low efficiency, prevents sporozoite invasion primarily through antibodies against the CSP protein, a significant component of the sporozoite pellicle.
In animals and humans, sterile protection against malaria can be obtained through immunisation with radiation-attenuated sporozoites, genetically attenuated sporozoites, or chemoprophylaxis with infectious sporozoites, but sporozoite-based live vaccines face significant challenges in terms of deployment. Antibodies against the sporozoite and CD8+ T lymphocytes against peptides presented by MHC class I molecules in infected hepatocytes mediate the protection induced by sporozoites in the pre-erythrocytic phases. As a result, the discovery of malaria antigens expressed in the sporozoite and liver stages may lead to the development of new vaccine candidates that can be used as recombinant protein-based virus-like particles or sub-unit virallyvectored vaccines, either alone or in combination [1].
Plasmodium sp. are polymorphic, obligate intracellular parasites with a complex life cycle that includes both an asexual and sexual stage, and parasitize two very different hosts: invertebrates, primarily Anopheles mosquitos, and vertebrate mammals. Although infections in other primate species have been observed with these pathogens, the five species P. falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi are recognised as the cause of natural infection in humans [2].
Malaria is spread by Plasmodium-infected Anopheles female mosquitos of various species. During a blood meal, an infected female mosquito injects roughly one hundred sporozoites into the skin of a mammalian host. The sporozoites look for capillaries in the epidermis and penetrate the host's circulatory system within minutes, eventually infecting liver cells. The sporozoites in the liver pass through multiple hepatocytes before successfully infecting one and dwelling in a parasitophorous vacuole with unique functions. For P. falciparum, the hepatic infection is asymptomatic and takes around 7 days to complete. P. vivax and P. ovale can develop a latent form, the hypnozoite, which can stay in the liver and trigger relapses months or years later by infecting the bloodstream. Parasites generate tens of thousands of merozoites in hepatocytes after undergoing asexual schizogony. Exoerythrocytic forms are parasites that live inside the hepatocyte (EEFs). Both the sporozoite invasion of the mammalian host and the liver-stage are included in the pre-erythrocytic stage, also known as the exo-erythrocytic cycle. This stage of the disease is appropriate for chemotherapeutic and immune prophylactic therapies to prevent the disease from progressing to clinical malaria [3].
Following merozoite egress from the infected liver cell, the parasites enter the bloodstream and infect erythrocytes, where they reproduce asexually. The symptomatic and well-studied blood-stage malaria, or erythrocytic cycle, is this. Merozoites infect erythrocytes and transform into trophozoites, which then grow into schizonts, which burst the host cell and infect new erythrocytes. Some trophozoites, on the other hand, grow into gametocytes. When a female mosquito consumes a blood meal from an infected mammalian host that contains at least one female and one male gametocyte, the parasite resumes sexual development. The sexual stage, or sporogonic cycle, takes place inside the mosquito, resulting in gametes that mature into ookinetes, which then pass the mosquito's midgut wall to produce oocysts. Thousands of sporozoites are generated inside oocysts [4].
Malaria-related immune response
Molecular components of the innate and adaptive immune systems have a role in disease aetiology and regulation. Clinical immunity to malaria can be gained in three stages: illness immunity, symptomatic infection immunity, and partial parasitemia immunity. Premunition (lack of fever with infection and reduced parasitemia density) is found in malaria-endemic areas and in people who have had multiple infections over the years (8–15 years), resulting in innate immune responses that reduce the likelihood of clinical illness. During epidemiological research with individuals from endemic locations who can regulate parasitemia and acquire a subclinical illness in the early 1900s, the term was coined. It has a sluggish acquisition rate, is only found in holo- or hyper-endemic locations, is quickly lost, is strain dependent, IgG dependent, and is directed towards blood-stage parasites; while the immune response elicited is powerful, it is not sterilising immunity. The mechanism of protection has not been fully explained, although evidence suggests that cytophilic antibodies and memory cells created after recurrent infections with Plasmodium variations are involved [5].
During Plasmodium infection, an innate immune response is generated as the first line of defence, followed by an adaptive immunological response that comprises T-cells, B-cells, and antibodies. When a mosquito bites a person, it injects Spz into their skin, which can stay there for up to 6 hours. The site of antigen presentation, as well as the location and type of response elicited, are all affected by this retention.
RBC haemoglobin is the parasite's primary source of protein. Hydrolysis of haemoglobin releases heme, a lipophilic prosthetic group that is particularly poisonous to the parasite. Heme detoxification is consequently required, which is accomplished by converting heme to hemozoin (Hz), an insoluble crystalline substance. In the case of P. falciparum infection, Hz binds DNA inside the phagolysosomes and cytosol of the host cell, and nucleic acids, NLRP3, AIM2, and other cytosolic sensors activate TLR9.
During the hepatic stage, antigens are exposed on hepatocyte surfaces in conjunction with MHC class I molecules expressed on all nucleated cells in order for CD8+ T-cells to detect them. Immunoglobulins can protect or stop disease progression in a variety of ways, including blocking Spz from entering hepatocytes with neutralising anti-Spz antibodies. Specific antibodies that trigger cell-mediated death or prevent RBC invasion and block the proteins responsible for binding to substances on the cell surface can opsonize Mrz in the erythrocyte stage.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
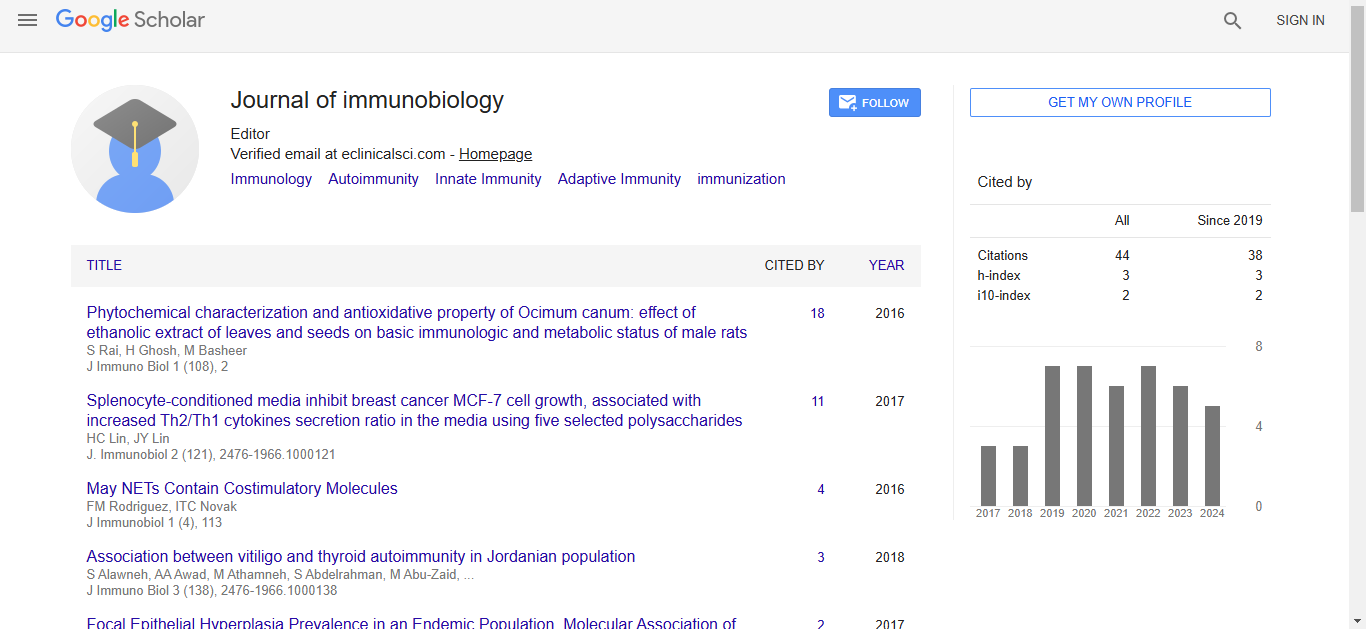
Journal of Immunobiology received 34 citations as per Google Scholar report