Review Article - (2021) Volume 9, Issue 7
Received: 11-Oct-2020
Published:
27-Jul-2021
, DOI: 10.37421/2329-9517.2021.9.459
Citation: Shivangi Chauhan. “Relation between Mitochondrial Dysfunction and Cardiovascular Disease". J Cardiovasc Dis Diagn 9 (2021) 459
Copyright: © 2021 Chauhan S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Globally, cardiovascular disease (CVD) is considered to be an important cause of mortality. The proper functioning of mitochondria is essential in tissues and organs which require high energy, involving the heart. Mitochondria are extremely sensitive towards nutrient and oxygen supply and experience metabolic adaptation due to the change in environment. Such adaptation is damaged in CVD that sequentially results in the degeneration of mitochondrial function correlated with irregularities in the respiratory chain and ATP production, raised oxidative stress, and loss of the structural integrity of mitochondria. Uncoupling of the respiratory chain in dysfunctional mitochondria leads to higher production of reactive oxygen species(ROS), reduction of cell ATP pool, massive cell damage, and apoptosis of cardiomyocytes. In a mitophagy process, cells clear themselves from impaired and dysfunctional mitochondria by autophagic mechanism. During heart failure, this process gets deregulated which leads to the accumulation of dysfunctional mitochondria and the situation became more adverse. Abnormalities of the respiratory chain activity and ATP synthesis may be reflected as a core of mitochondrial dysfunction in cardiac pathology. Indeed, the main objective for improvement of mitochondrial dysfunction in CVD is to therapeutically restore these vital functional properties.
Mitochondrial dysfunction; Cardiovascular disease; ATP reduction; Cell apoptosis; Oxidative stress; Autophagy
Cardiovascular disease (CVD) which is extensively distributed in the world signifies a major cause of mortality in humans. The annual (2017) report of the American Heart Association (AHA) represent that 92.1 million adults in America were affected with some form of CVD. CVD which is the featuring source of death causes around 801,000 deaths annually, which is nearly 1/3 (31%) of the global mortality in the US. The most common cause of CVD-induced mortality is Coronary artery disease (CAD) accounting for 45% of cases [1]. In the US, the population ageing and obesity is principally attributed for the growth of CVD incidence.
Mitochondria are cellular organelles that originate maternally and are responsible for the production of energy via the process of oxidative phosphorylation. In the production of energy numerous organic substrates are utilized, usually fatty acids and less commonly glucose [2].
Mitochondria are considered to be the site for ROS production during the enzymatic activity of electron transport chain [3]. The etiology of CVD is very complex. Several risk factors and pathological mechanisms are responsible for the influence of this disease. In cells, excessive ROS production and impairment of mitochondrial dynamics lead to mitochondrial dysfunction and ultra-structural changes of cellular lipids, proteins, enzymes and DNA which in turn results in the development of several cardiac diseases [4]. Additionally, mitochondria may lead to regulation of cell stress response by retrograde signalling, as soon as lack of the mitochondrial membrane potential causes activation of a number of signalling proteins, which, in turn, results in the upregulation of many stress-responsive genes in the nucleus [5]. This describes the complicated clinical manifestations exposed in mitochondrial disorders usually appeared in tissues that require high energy supply, such as skeletal muscles, heart, and kidney [6]. Thus, aiming of mitochondrial dysfunction is an essential step in the treatment of a variety of cardiac diseases and several approaches have been tested in experimental and clinical studies with, however, controversial findings. This review studied the modifications of the mitochondrial function which influences CVD pathogenesis, with emphasis to specific CVD forms.
Morphology of mitochondria in a stable myocardium
Mitochondrion is a double membrane cellular organelle of size generally varies from 0.75 to 8 μm [7]. In order to meet the increased energy requirements, the cardiac cells contain vast amount of mitochondria which produced approximately 6kg of ATP daily through the oxidative phosphorylation process [8]. There are two mitochondrial populations i.e., interfibrillar and subsarcolemmal mitochondria organized in cardiomyocytes. There is a formation of parallel series along the length in Interfibrillar mitochondria and the orientation of their cristae is mostly tubular. The clusters of Subsarcolemmal mitochondria are organized beneath the sarcolemma and show lamellar cristae orientation [9]. In cardiac muscle cells, both mitochondrial populations are electrically attached to each other supplying electric conduction from one mitochondrion to another [10]. The fusion and fission proteins control the mitochondrial biogenesis and morphology and are responsive to alterations in cardiac muscle cells [11]. Moreover, morphology of mitochondria is dynamic and sensitive to cardiac alterations involving those that occur in heart pathology [12].
Morphology of mitochondria within cardiac pathology
Association of mitochondrial structure modifications in the tissue with cardiac pathologies involves the development of megamitochondria (also called giant mitochondria) of size upto 30 μm [13]. Megamitochondria are produced through fusion of adjacent organelles as a result of overexpression of fusion proteins, such as mitofusin 1 (Mfn1) and Mfn2 [14] or through enlargement of an individual mitochondria. Megamitochondria did not have any functional utility and abolished as a result of mitophagy mechanism [12]. Other pathological changes in the structure of mitochondria includes loss or reorientation of cristae, formation of intra-mitochondrial rods and crystalloids mainly composed by creatine kinase crystals [15]. Ischemic Heart causes fragmentation of mitochondria owing to up-regulation of dynamin-related protein 1 (Dnp1) which is a fission protein [16]. This protein is able to form a proapoptotic complex with Mfn2 and BAX on the outer mitochondrial membrane which is accompanied with opening of the mitochondrial permeability transition pore (MPTP), release of cytochrome c, fragmentation of mitochondria, and initiation of cardiomyocyte apoptosis [17]. Dnp1 is inhibited for the protection of cardiac mitochondria from fragmentation and prevents apoptosis [16].
Dysfunctional mitochondria and CVD
In CVD, the Functional abnormalities of cardiac mitochondria results in increased oxidative stress, reduced ATP generation and energy supply, raised cell apoptosis, and impaired mechanisms of autophagy [18].
Excessive production of ROS in CVD: The mitochondrial respiratory chain is considered to be the leading pathway in the production of energy stored in ATP molecules. In normal conditions, the respiratory chain facilitates the production of ATP using more than 98% of the electron transport. Only 1-2% of electrons are discharged for the generation of superoxide radical which, in turn, is decomposed by superoxide dismutase [19]. The regulation of generated ROS is significantly involved in the initiation of protective signaling mechanisms in ischemic preconditioning [20]. Excessive production of ROS is due to the uncoupling of the mitochondrial electron transport chain from ATP generation, which then causes oxidation of lipids and proteins and extensive cell destruction. Excess production of ROS encourages atherogenesis at all stages by inducing endothelial dysfunction, vessel inflammation, accumulation of oxidized low dense lipoprotein (oxLDL) in the arterial wall, formation of the initial lesion and its maturation to the advanced plaque with possible progression to plaque rupture [21].
Reduction in ATP generation in CVD: The electron transport chain located in the internal membrane of mitochondria accomplished the synthesis of cellular ATP via OXPHOS mechanism [22]. The ATP levels in the heart are relatively low because of exhaustive ATP intake and fast ATP turnover [23]. The creatinine kinase present in the mitochondria is engaged in monitoring the activity of the respiratory chain and integrating the high-energy metabolism of phosphate. This enzyme initiates the production of phosphocreatine from creatine which primarily helps in maintaining the ATP buffer content of the myocardium [24]. The two forms of enzyme exist: highly-reactive octameric and less active dimeric which present in dynamic balance. During heart disease, as the dissociation of the octamer increases the equilibrium between both forms is shifted towards the dimer [25] which leads to the formation of inactive crystalloids from the octameric creatine kinase [15]. Consecutively, this shows impairment in the respiratory control and compensation of high cardiac ATP depletion by phosphocreatine hydrolysis. Damaged OXPHOS and decreased ATP levels existing in the failing heart as compared to the normal heart results in reduced cardiac performance [26]. In an adult heart 50-70% of energy is attained by mitochondrial β-oxidation of fatty acids [27]. Depending on the availability of nutrient, the myocardium dynamically switched from the preferential use of lipids to glucose as the energy supply to maintain the constant ATP production. Insulin dependent signaling regulates this substrate selection flexibility in cardiomyocytes [28]. On the other hand, in failing heart, there is a down-regulation of insulin signalling due to which the metabolic flexibility gets damaged. Hence, the reduction of fatty acid oxidation is unable to compensate by an increase in glucose oxidation as an alternative source of energy which results in the depletion of cardiac ATP [29].
Enhanced apoptosis within CVD: The inherent mechanism of apoptosis of cardiomyocytes is initiated by dysfunctional mitochondria in case of reduction in respiratory chain activity, ATP depletion and enhanced oxidative stress [30]. Irretrievable damage of cell produced by acute or chronic ischemia and metabolic stress causes the introduction of apoptosis. As stated above, the apoptotic death of cardiac muscle cells is associated with ischemia/reperfusion injury [31] through initiation of mitochondrial fragmentation, formation of the pro-apoptotic assembly between Dnp1, Mfn2, and BAX, enhanced mitochondrial permeability and the release of cytochrome c to the cytosol [17]. The multiprotein complex is formed by cytochrome c with apoptotic proteaseactivating factor 1 (APAF1), which later stimulates procaspase-9. As soon as it gets stimulated, the effector caspases is activated by the initiator caspase that trigger a cascade of irreversible actions directing to apoptosis [32]. In cardiac ischemia, mitochondria released a mitochondrial serine protease known as high temperature requirement A2 (HtrA2) into the cytoplasm, where it participates in the activation of caspase, thereby promoting apoptosis [33]. The cardiomyocytes apoptosis is controlled by pro-apoptotic BH3 domainonly factors such as heart-expressed BNip3 and Nix (also known as BNip3L). Heterodimers were formed from these proteins with anti-apoptotic factors such as Bcl2 and BclX1 which further allows the activation of pro-apoptotic proteins BAX and BAK. Nix also causes programmed necrosis of cardiac muscle cells through the stimulation of Ca2+ released from the sarcoplasmic reticulum. MPTP opening is initiated due to the rise of Ca2+ in cytosol resulted in the depletion of ATP and cell death [34]. However, in the ischemic myocardium, the initiation of cardiomyocytes apoptosis requires BNip3 whereas in hypertrophied myocardium the cardiomyocytes death was facilitated by Nix [35]. Indeed, these factors are essential in ischemia- and non-ischemiainduced cardiac alteration, while apoptotic and necrotic cardiomyocytes are exchanged by fibroblasts in progression of heart scarring and cardiac fibrosis [36]. Hence, cardiac stress prompted mitochondrial dysfunction which later activates subsequent apoptosis of cardiomyocytes.
Deregulation of autophagy during CVD: The mitophagy that are accountable to clear out the dysfunctional mitochondria is constantly functioned in normal cardiomyocytes to support cell survival by eradicating longlived organelles and proteins [37]. In moderate cardiac stress, mitophagy is able to effectively remove the dysfunctional mitochondria. Mechanically, stress encourages the growth of PTEN induced putative kinase 1 (PINK1) on the surface of mitochondria. PINK1 phosphorylates Mfn2 in order to interact with Parkin which is an E3 ubiquitin ligase [38]. Parkin ubiquitinates main mitochondria-associated proteins can be then recognized by autophagy adaptor proteins. The recruitment of autophagosomes through autophagy adaptors to targeted mitochondria were followed by autophagosomalmitochondrial fusion and degradation [39].
The autophagy flux declined because of the damaging events such as acute cardiac ischemia-reperfusion injury. As a result, there is an accumulation of damaged mitochondria in cardiomyocytes that leads to severe oxidative stress and causes cardiomyocyte apoptotic death [40]. was shown to be involved in the pathogenesis of various CVDs, including ischemic heart disease, cardiac hypertrophy, heart failure, and dilated cardiomyopathy implicated the loss of a proper autophagy mechanism.
Dysfunctional mitochondria in certain CVD forms
Atherosclerosis: Mitochondrial dysfunction leads to excessive oxidative stress which is an important pro-atherogenic molecular mechanism. However, oxidative metabolism seems to be of less significance for vascular endothelial cells (ECs), where the major energy source is a glycolytic process [41]. In spite of that, mitochondria are crucially involved in EC functioning as initiate production of nitric acid (NO), apoptosis, intracellular signaling, etc. [42]. Excess production of ROS was responsible to stimulate EC senescence [43], in order to induce apoptosis and promote atherogenesis [44]. In vascular smooth muscle cells (VSMC), the mitochondrial function is inhibited by oxLDL with the down-regulation of the respiratory activity and ATP generation and increases VSMC hyperplasia, migration, and proliferation, specifically promotes the formation of pro-atherogenic neointima [45]. Interestingly, the proliferation of VSMCs were blocked by hyperplasia suppressor gene (HSG), a rodent ortholog of human fusion protein Mfn2. Studies in apolipoprotein E (apoE)-deficient hypercholesterolemic mice, an atherosclerotic model have demonstrated a great reduction in HSG expression in affected arteries, thus enhanced the progression of atherosclerosis and proliferation of vascular smooth muscle cells (VSMCs) [46,47]. Similarly a study in rabbit as a model of atherosclerosis demonstrated the anti-atherogenic properties of human Mfn2, when excess Mfn2 production leads to reduced VSMC proliferation/ hyperplasia and diminished progression of plaque [48]. Therefore, mitochondrial dysfunction is responsible to initiate as well as progress the atherosclerotic vascular disease by exerting opposite effects on vascular cells and induce senescence and death of ECs. So, it can be considered as an important target for the treatment of CAD.
Ischemic heart disease: The mitochondrial OXPHOS is responsible to compensate the high demand of energy in cardiac cells. Indeed, cardiomyocytes are extremely susceptible to oxygen deficiency. In order to evaluate the effects of oxidative stress on cardiomyocytes, ischaemia is considered to be the best experimental model. In heart ischemia, cardiomyocytes become hypoxic and induce mitochondrial dysfunction. Ischemia/reperfusion has damaging effects on mitochondrial function by enhancing ROS production and increasing Ca2+ flux into mitochondria. As a result the electrochemical gradient across the inner mitochondrial membrane fails and deteriorate the activity of mitochondrial respiratory chain enzymes [49,50]. Damaged mitochondrial cardiolipin is associated with the reduced functional capacity of Complex-I activity which promotes electron leakage and accelerates a vicious cycle of production of free radicals [51].
During ischemia, early initiation of autophagy was shown as an adaptive response to cardiac injury [52]. The up-regulated autophagy markers can moderately reflect either elevation or insufficiency of autophagy flux due to the damage of strict control [53]. There are adverse effects of excessive autophagy in acute cardiac injury as it leads to deprivation of essential and fully functional organelles and proteins. However, autophagy is inhibited at late stages of heart disease [54], and fewer numbers of autophagic vacuoles can be linked with poor diagnosis for diseased patients [55].
Cardiac hypertrophy and Heart failure: In heart failure, structural and/or functional cardiac abnormalities are involved which result in degraded cardiac output and/or raised intra-cardiac pressures at rest or during stress [56,57]. Cardiac hypertrophy is the early response to increased cardiac workload which is termed as the thickening of ventricular wall and decline in ventricular volume [58]. In cardiac hypertrophy, excess energy is required for the growth of cardiomyocyte and thus should be associated with increase in mitochondrial abundance. In hypertrophied rat hearts, mitochondrial enlargement was observed after aortic constriction [59].
However, during cardiac hypertrophy, at early disease stage it was discovered that the mitochondrial count increased initially and degenerates with the advancement of disease that is frequently associated with contractile abnormalities [60]. As a consequence, ability of mitochondria is reduced to up-regulate ATP synthesis and causes reduced cardiac relaxation followed by diastolic dysfunction and heart failure [61].
The growth of cardiac hypertrophy, remodeling and HF are associated with increased production of ROS and damaged function of the anti-oxidant systems [62,63]. In heart failure, membrane rupture and matrix depletion leads to impairment in mitochondria [64]. As a result of the impaired activity of the respiratory chain and reduction in OXPHOS capacity, these mitochondria demonstrate inadequate capacity for the production of ATP [65,66]. In heart failure subjects, abnormalities in the respiratory chain were associated with reduced activity of complexes I and IV [67]. Actions of other redox enzyme, NADPH-transhydrogenase, and the Krebs cycle enzyme such as isocitrate dehydrogenase, malate dehydrogenase, and aconitase are intensely reduced during end- stage heart failure. The abundant chemical modification leads to reduced activities of mitochondrial enzymes [68]. In addition, in human failing hearts, it was observed a significant reduction in the copies of mtDNA, which results in mtDNA exhaustion, prominent decrease of mtDNA-encoded proteins, and finally disruption of mitochondrial biogenesis [69,70]. In conclusion, it was suggested that mitochondrial dysfunction may play crucial role in the induction and pathogenesis of heart failure due to the essential role of mitochondria in the production of energy [71].
The function of mitochondria is essential role in all human cells and tissues/organs especially in heart and others which require higher energy. Mitochondria produce energy through oxidative phosphorylation and hence the activity of the electron transfer chain and ATP-producing machinery has to be firmly regulated. Additionally, mitochondria are cellular organelles whose morphology and cellular count are able to vigorously change in response to outer signals. The balance between mitochondrial fission and fusion as well as mitochondrial biogenesis should be also accurately controlled. Several CVD such as atherosclerosis, ischemia-reperfusion injury cardiac hypertrophy and heart failure are associated with abnormal mitochondrial function and excessive ROS production which damage cellular lipids, proteins, enzymes and DNA. For example, in ischemia/reperfusion injury, the balance towards fission is shifted due to increased oxidative stress and Ca2+ flux, which consecutively results in mitochondrial fragmentation and activates apoptosis. If it results in fission overactivation, it can be also lethal in heart failure and diabetic hyperglycemia. In the heart, the main source of energy are mitochondria, thus any abnormalities in the activity of the respiratory chain and ATP synthesis may be reflected as a core of mitochondrial dysfunction that then causes induction of oxidative stress, apoptosis, abnormal autophagy and other pathophysiological perturbations examined in CVD. Therefore, it is advised to restore the respiratory activity and ATP-generating capacity of mitochondria as a primary therapeutic target in order to recover the mitochondrial dysfunction in various CVD forms.
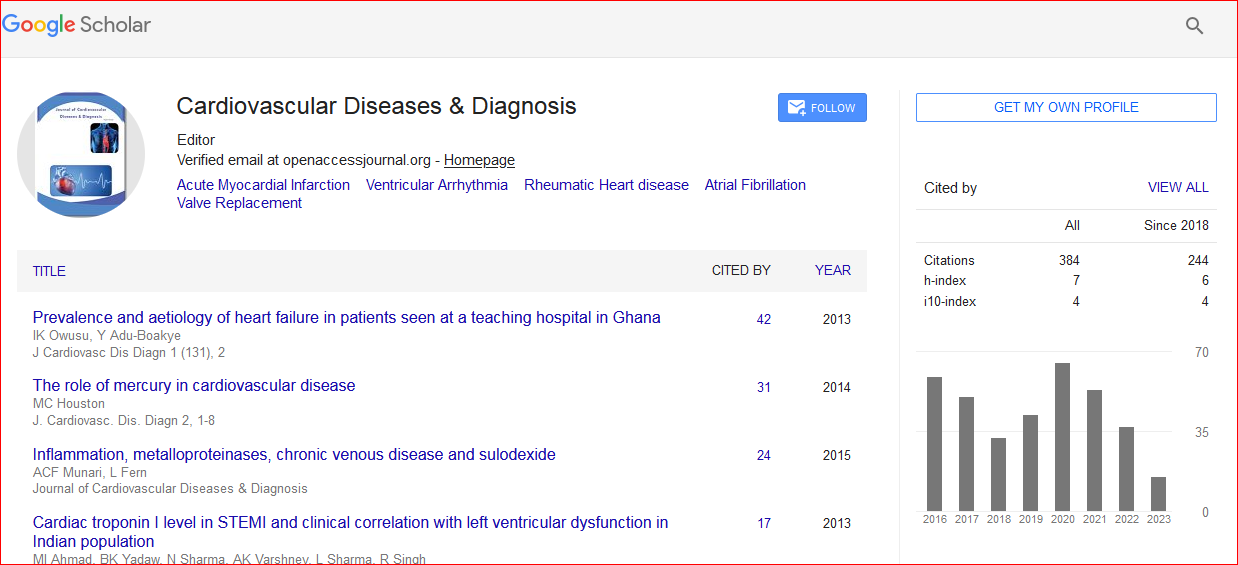
Cardiovascular Diseases & Diagnosis received 427 citations as per Google Scholar report