Mini-Review - (2022) Volume 13, Issue 11
Received: 02-Nov-2022, Manuscript No. jch-22-84337;
Editor assigned: 05-Nov-2022, Pre QC No. P-84337;
Reviewed: 16-Nov-2022, QC No. Q-84337;
Revised: 22-Nov-2022, Manuscript No. R-84337;
Published:
28-Nov-2022
, DOI: 10.37421/2157-7099.2022.13.662
Citation: Luongo, Marianna. “Ribosome Synthesis and Translational Ability in Comparison to Cell Growth." J Cytol Histol 13 (2022): 662.
Copyright: © 2022 Luongo M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Ribosome and protein synthesis are central to cell growth and major users of the cellular budget.In this paper, we discuss recent advances in the coupling of ribosome synthesis and translational capacity with bacterial cell growth.We discuss the various strategies used by bacteria to modulate protein synthesis rates at fast and slow growth rates.Bacterial cells,in particular,maintain translational potential at very slow growth as a strategy for maintaining fitness in changing environments.We also discuss the critical role of ribosome synthesis in rapidly proliferating eukaryotic cells like yeast and cancer cells.The close relationship between ribosomes and cell growth opens up a wide range of research opportunities for scientists from various disciplines.'The dream of every cell is to become two cells,' said Francois Jacob.Growth is a basic characteristic of many cell types, including bacterial cells,yeast cells,and cancer cells.Bacterial cells rarely encounter stable nutrient-rich conditions in their natural niche, instead frequently undergoing feast-and-famine cycles (see Glossary),and this process can be mimicked in the laboratory by varying the combinations of nutrient sources.Understanding how bacterial cells adapt to different environments, on the other hand, remains a major challenge in biology.
Cells • Bacterial • Yeast • Ribosome
Under different nutrient conditions, the cellular ribosome content of E. coli cells increases linearly with growth rate, which is known as 'growth-rate control of ribosome synthesis.' This phenomenon reflects the concern about bacterial cells' optimal proteome resource allocation in response to nutrient availability. The synthesis of ribosomes (including rRNA and r-proteins) and other translation-associated proteins is critical in the cellular budgeting process because it consumes a significant amount of cellular resources. Meanwhile, to support growth, cells must devote a significant portion of their resources to metabolic proteins, which include both catabolic and anabolic proteins that convert external nutrients into the raw materials, 20 amino acids, for protein translation.To maximise growth rate, bacteria must maintain the balance between ribosomes and metabolic proteins.
When nutrient sources are plentiful, the cellular budget of metabolic proteins is low because amino acids and other growth factors are already available for use, allowing bacteria to maximise ribosome synthesis in order to achieve rapid growth. However, when nutrient sources become scarce (for example, in a glycerol minimal medium with a DT of 60 minutes), cells must synthesise a large amount of metabolic protein for nutrient uptake and processing at the expense of ribosomes, resulting in a slower growth rate. The recently developed bacterial 'growth law' model of proteome resource allocation quantitatively accounts for the trade-off between ribosomes and metabolic proteins, as well as their links to cell growth [1].
The alarmone guanosine tetra- or pentaphosphate (also known as the'magic spot') regulates the proteome resource allocation between ribosomes and metabolic protein;ppGpp is a global stress regulator of bacterial cells. Its synthesis, mediated by RelA or SpoT, increases dramatically in response to nutrient deprivation and other types of stress, shutting down various biochemical processes such as replication, transcription, and translation; this is known as the stringent response. It also plays a role in the regulation of several critical physiological processes, including persistence, survival, motility, and pathogenesis.
Because ribosome synthesis is limited by rRNA synthesis and r-proteins are coordinatedly synthesised with rRNA via autogenous feedback control, regulating ppGpp on rRNA synthesis also allows it to control rprotein synthesis. As a result, when nutrients are scarce, the increased ppGpp pool inhibits ribosome synthesis, allowing E. coli cells to devote more proteome resources to metabolic proteins. Maintaining the balance between ribosomes and metabolic proteins in bacterial cells requires an optimal ppGpp level. Cell growth is inhibited when the cellular ppGpp pool is misregulated, owing to inefficient resource allocation among ribosomes and metabolic proteins. Furthermore, the ppGpp-null strain lacks growth-rate control of ribosome synthesis [2].
The abundance of tRNA ternary complexes (TCs), which is co-regulated with ribosome content by ppGpp, affects the elongation rate of ribosomes under different nutrient conditions. Because the charged fraction of tRNA does not change significantly under poor growth conditions, changes in amino acid concentrations are unlikely to affect translational elongation rate. Although the translational elongation rate decreases with poor nutrients due to lower TC levels, it remains significant at 8-9 aa/s during extremely slow growth or nutrient deprivation because TC levels remain significant under these conditions [3,4].
The precise mechanism by which TOR detects nutrient status is only partially understood at the moment. TORC1 nutrient sensors have been identified in S. cerevisiae as the small GTPase Gtr1/2 and the AMP protein kinase Snf1. Gtr1/2 regulates TORC1 by sensing the quality of cellular nitrogen and amino acids. TORC1 is a negative regulator of the AMPK protein Snf1, which is a glucose/energy sensor. In response to low cellular energy (high AMP:ATP ratio) and glucose deprivation, AMPK is activated. Mammalian cells have TOR signalling pathway regulatory mechanisms that are similar to human cells. Cancer is a broad term that encompasses a wide range of diseases characterised by key characteristics such as a high mutation rate, uncontrollable proliferation,evasion of growth suppressors, resistance to apoptosis, enhanced angiogenesis, immune evasion, invasion, and metastasis. Despite the complexities of cancer, all types of cancer have one thing in common: rapid and uncontrollable cell proliferation, which is similar to bacteria and yeast cells [5].
Ribosome synthesis is a critical process that drives protein synthesis and cell growth in many types of rapidly proliferating cells. Recent systematic studies have revealed the quantitative principles governing the interaction of ribosome synthesis and translational capacity with cell growth in microbial cells such as bacteria and yeast. Meanwhile, recent research has confirmed the importance of ribosomes in cancer progression. Despite their evolutionary distance and vast differences in gene regulatory networks, these rapidly proliferating cell types share some fundamental growth control design principles. Many important questions remain unanswered to this day, but several lines of research will provide valuable insight into the interdependence of ribosome synthesis, protein synthesis, and cell growth.
Such as ribosome profiling, will be useful in elucidating more aspects of the influence of ribosome abundance and translational efficiency on cell growth. These various types of proliferating cells (bacteria, yeast, and cancer) will hopefully bring together researchers from various disciplines to reveal more general growth control strategies. Acknowledgments We would like to thank members of the Terry Hwa laboratory for helpful discussions at various stages of this work.
None.
There are no conflicts of interest by author.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
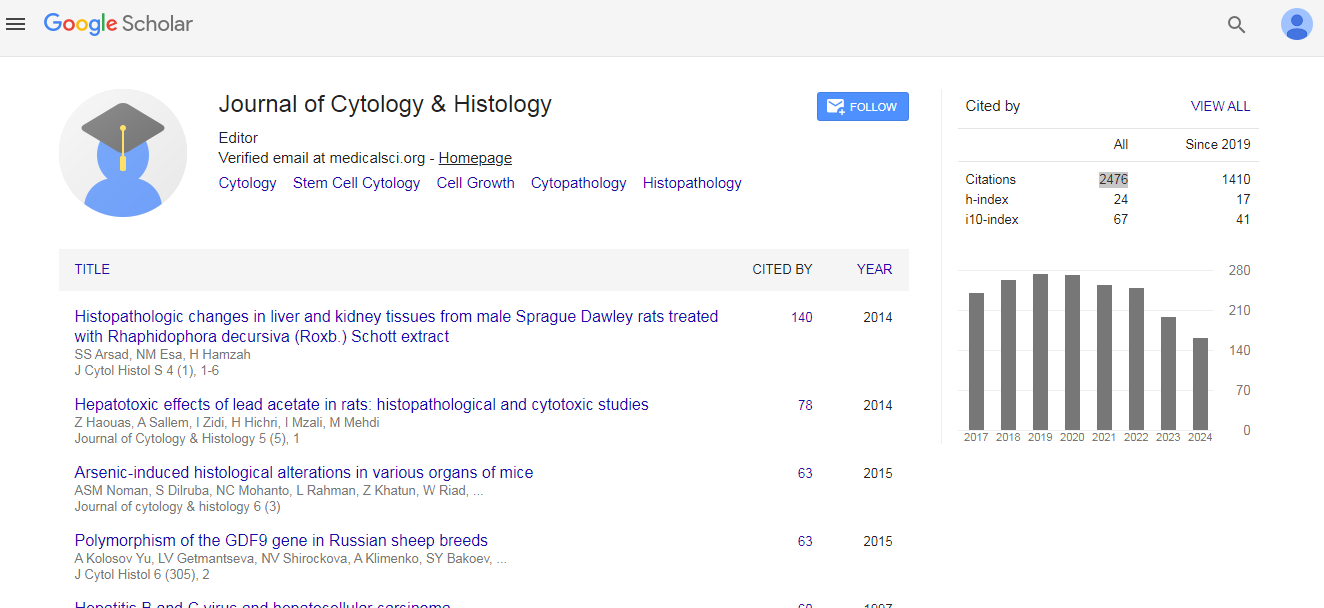
Journal of Cytology & Histology received 2476 citations as per Google Scholar report