Review Article - (2024) Volume 10, Issue 3
Received: 01-Jan-2025
Editor assigned: 04-Jan-2025
Reviewed: 18-Jan-2025
Revised: 22-Jan-2025
Published:
29-Jan-2025
, DOI: 10.37421/2572-4134.2024.10.229
Citation: Fukui, Hiroshi. "The Gut affects Pathophysiology
and Management of Diabetes Mellitus Type 2: The Recent Messages from
Intestine and Gastrointestinal Microbiota." J Food Ind Microbiol9(2025): 129.
Copyright: �?�© 2025 Fukui H. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author
and source are credited.
The number of diabetic patients rapidly increased in the world constituting a serious health problem. Daily life alterations such as high-fat high-sugar diet and insufficient physical exercise augment diabetes risk. Long-lasting low-grade inflammatory reactions in obese patients with metabolic syndrome are considered as an important role in the occurrence and advance in the stage of type 2 diabetes. Emerging clinical and experimental evidence reveals that gut dysbiosis, intestinal barrier disturbance and following metabolic endotoxemia are firmly related to the low-grade inflammatory, insulin resistance and resultant cardiovascular complications in patients with type 2 diabetes. Gut microbiome transmitted from mother to child at birth is deeply affected by dietary habits in life thereafter. In the feces of type 2 diabetics, relatively frequent abundance in endotoxin producing gram-negative bacteria and lower abundance in butyrate-producing bacteria were noted. Butyrate as an important energy source and a protector of intestinal barrier, its defect is considered to enhance intestinal hyperpermeability and metabolic endotoxemia. Inflammation in the adipose tissue induces detrimental effects on other organs and tissues through secreted pro-inflammatory cytokines. Activation of Toll-like receptor 4 in immune cells such as macrophages evokes inflammation and insulin resistance, finally leading to an impairment of insulin signal and β-cell failure. Inflammatory changes in the arterial vessels and liver lead to two life-threatening conditions, ischemic heart disease and liver cirrhosis, respectively. Careful management strategies to improve gut dysbiosis may stimulate effective drug treatment and lower the morbidity and mortality of patients who suffers from type 2 diabetes. A kind of biguanide metformin is considered as an anti-inflammatory influence in addition to its glucoregulatory effect. Two newly developed diabetic drugs, Dipeptydil-Peptidase-4 (DPP-4) inhibitors and Sodium-Glucose Co-Transporter 2 (SGLT2) inhibitors may in combination with reasonable dietary therapy have some undetermined effect on inflammatory changes. This systemic review summarizes the bulk of latest information published until 2021 on the pathogenesis and treatment of type 2 diabetes, especially related to gut microbial alterations. This manuscript in addition to my previous review includes up-to-date information in diabetes mellitus type 2.
Type 2 diabetes • Intestinal permeability • Gut dysbiosis • Endotoxemia • Inflammation • Prognosis • Dietary therapy • Metformin • Dipeptydil-peptidase-4 (DPP-4) inhibitors • Sodium-glucose cotransporter (SGLT)-2 inhibitors
The number of type 2 diabetic patients is increasing in the world these years. The Diabetes affects an estimated 34 million US adults, in 2020 [1,2]. In addition to genetic predisposition, this remarkable increase in diabetic patients should be caused by changes in lifestyle, i.e. diet, nutritional condition and physical activity [3-5]. It is now evident that the advanced pancreatic β-cell failure that induces the deterioration of metabolic control over time begins early and is existing long before the diagnosis of diabetes [6]. Obesity and pre-diabetes as underlying risk factors predisposing type 2 diabetes and related complications is recommended to be targeted in the diabetes prevention program. Although lifestyle adjustment is basically indicated for every overweight and obese patient with prediabetes, useful pharmacotherapy can be developed simultaneously for preserving β-cell function in type 2 diabetics [7].
The potential role of the intestinal epithelial barrier dysfunction and increased permeability has been described these days associated with gut dysbiosis in various human diseases. The intestinal diseases including inflammatory bowel disease and irritable bowel syndrome are related to gut dysbiosis [8-15]. Additionally, the relations of liver, pancreas, kidney, heart and brain diseases together with systemic autoimmune and allergic diseases to gut microbiota have been described as well [16-37]. Among them, Type 2 diabetes and gut dysbiosis should be counted as one of popular entities. The gut barrier derangement connected with intestinal dysbiosis is possible to induce the gut translocation of fragments originated from microbiome and the development of "metabolic endotoxemia", inducing systemic low-grade inflammation and insulin resistance [38].
Adipocytes are massive sources of bioactive substances named adipocytokines (adipokines) [39]. Adipokine dysregulation due to endotoxemia and excessive fatty acids possibly cause obesity-related metabolic disorders, called metabolic syndrome, a cluster of metabolic disturbances, including type 2 diabetes, hyperlipidemia, hypertension, Nonalcoholic Fatty Liver Disease (NAFLD) and Nonalcoholic Steatohepatitis (NASH) [39,40]. The imbalance between excessive pro-inflammatory cytokines including Tumor Necrosis Factor-α (TNF-α) and defective anti- inflammatory adipokines including adiponectin is an important etiologic background in the adipose tissue from fatty patients with metabolic syndrome [41].
The etiology of type 2 diabetes is firmly related to the intestine. The fundamental lifestyle care begins with dietary therapy, which should be drawn from the science of gut microbiota, because diet itself profoundly affects gut microbiome and finally gut dysbiosis. The effect of diabetic therapy is fundamentally bound by basic dietary care. As described in the abstract, this review introduces latest knowledge of gut dysbiosis, intestinal hyperpermeability, endotoxemia and their cardinal meanings in the fundamental therapy of subjects with type 2 diabetes.
Gut microbiota, as a complex intestinal "superorganism", influences host metabolic state modulating appetite, energy absorption, gut motility, glucose and lipid metabolism, in addition to hepatic fatty storage [38]. Small Intestinal Bacterial Overgrowth (SIBO) is defined as a condition in which colonic bacteria move into the small intestine due to impaired microvilli function. This causes a breakdown in intestinal motility and resultant disturbed gut homeostasis [42,43]. Important examinations showed higher prevalence of small intestinal bacterial overgrowth (SIBO) (50%~77.8% vs. 9.1%~31.2%) in NAFLD patients compared with healthy controls [44–48]. SIBO was further reported to be related to enhance hepatic expression of Toll-Like Receptor (TLR) 4 and Interleukin (IL)-8 in patients with NASH [46].
Related to the above findings, SIBO was also noticed in 43% of diabetic patients complaining chronic diarrhea and 75% had a marked improvement in their symptoms after being treated with antibiotics [49]. In addition, a group of 82 diabetic patients with carbohydrate malabsorption on an oral glucose tolerance test, 75% were found to reveal SIBO [50,51].
Accumulating data in human studies and animal experiments suggest that obesity and type 2 diabetes are related to a severe gut dysbiosis [52]. Ley et al. first found in their preliminary report that obese patients had higher abundance of Firmicutes and lower abundance of Bacteroidetes compared with lean controls [53]. Turnbaugh et al. revealed that the microbial alterations affect the metabolic potential of the mouse gut microbiome in the way that the microbiota from an obese animal shows an increased capacity to absorb energy from the diet [54]. They noted that microbiota transplantation from obese mice fed a high-fat diet to lean germ-free counterparts promoted greater fat deposition than recipients from lean counterpart donors [55]. They further disclosed that the human gut ecosystem in mice by transplanting human fecal microbiota into germ-free mice and revealed that the high-fat, high-sugar diet altered microbiota gene expression [56]. These mice had increased adiposity and their microbiota showed an increased abundance of Erysipelotrichaceae class bacteria and that of the Bacilli (mainly Enterococcus) within the Firmicutes phylum in addition to a decreased abundance of members of the Bacteroidetes [56]. In contrast to these clear-cut experiment results, the following clinical studies on the gut microbiome have given various and often contradictory results as reviewed in the previous article [20]. Some studies even reported an increased Bacteroidetes/Firmicutes ratio in obese patients [57-59]. High dietary fructose was related to lower abundance of the beneficial microbes such as Eubacterium and Streptococcus, which involves carbohydrate metabolism [60]. Multiple factors, including unhealthy dietary habits, can cause disruption of microbiota equilibrium (dysbiosis) with extra-intestinal metabolic disturbances including obesity and type 2 diabetes [61].
In addition to bacterial cells and bacterial structural components, bacterial metabolites also work the health and disease of the host subjects [20]. Human colonic bacteria decompose substrates including resistant starch and non-starch polysaccharides as main components of dietary fiber, which are not completely hydrolyzed by host enzymes in the small intestine [62]. The major fermentation products from this fiber breakdown are the Short Chain Fatty Acids (SCFAs) such as acetate, propionate and butyrate [62].
Alterations in gut microbiota presented in recent clinical studies on type 2 diabetes are described in Table 1. As noted in the previous review, the results again seem surprisingly variable, while noteworthy common microbiota characters are there observed [63]. Larsen et al. described that feces in patients with type 2 diabetes were comparably enriched with endotoxin producing gram-negative microbiome, belonging to the phyla Bacteroidetes and Proteobacteria[64]. Chinese type 2 diabetic patients were also characterized by an evident gut microbial dysbiosis, a decrease in some universal butyrate-producing microbiome (Clostridiales sp. SS3/4, Eubacterium rectale, Faecalibacterium prausnitzii, Roseburia intestinalis and Roseburia inulinivorans etc.) and an increase in variable opportunistic pathogens, such as an increase in other microbial functions revealing sulphate reduction and oxidative stress resistance [65]. Another large-scale Swedish study also showed decrease in butyrate-producing bacteria like Roseburia and Eubacterium eligens in female type 2 diabetic patients.
| Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|
| Firmicutes↓ | Bacilli↑ | Lactobacilales | Lactobacillace↑ | Lactobacillus ↑ | L. gasseri ↑ |
| L. plantarum ↑ | |||||
| L. reuteri ↑ | |||||
| Aerococcaceae | Abiotrophia ↑ | ||||
| Streptococcaceae | Streptococcus ↓ | S. mutans ↑ | |||
| Enterococcaceae | Enterococcus | E. rectale ↓ | |||
| Clostridia↓ | Clostridiale↑ | Clostridiaceae | Clostridium ↓ | *Clostridiales ss3/4 ↓ ↑ | |
| C. coccoides ↓ | |||||
| C. hathewayi↑ | |||||
| C. ramosum↑ | |||||
| C. symbiosu↑ | |||||
| C. clostridioforme ↑ | |||||
| Sporobacter ↑ | |||||
| Subdoligranulum ↑ | |||||
| Peptostreptococcus ↑ | |||||
| Eubacterium ↑ | *E. rectale ↓ | ||||
| *E. eligens ↓ | |||||
| Ruminococcaceae | Ruminococcus ↑ | ||||
| Faecalibacterium | *F. prausnitzi ↓ | ||||
| Lachnospiraceae | *Roseburia | *R. intestinales ↓ | |||
| *R. inulinivoran ↓ | |||||
| *Roseburia ↓ | |||||
| Dorea ↑ | |||||
| Negativicutes | Salenomonadales | Veillonellaceae | Megamon ↓ | ||
| Actinobacteria | Coriobacteriaia | Coriobacteriales | Coriobacteriacea ↓ | Coriobacterium | Atopobium ↓ |
| Collinsella ↑ | |||||
| Eggerthellales | Eggerthellaceae | Eggerthella | E. lenta ↑ | ||
| Verrucomicrobia | Verrucomicrobiae↓ | Verrucomicrobiales | Verrucomicrobiaceae | Akkermansia | A. muciniphilia ↓ |
| Bacteroidetes | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides ↓ | B. intestinalis ↓ |
| B. caccae ↑ | |||||
| Prevotellaceae | Prevotella ↑ | ||||
| Proteobacteria | β-proteobacteria↑ | ||||
| γ-proteobacteria | Pasteurellales | Pasteurellaceae | Haemophilus ↓ | ||
| Enterobacteriales | Enterobacteriaceae | Escherichia | E. Coli ↑ | ||
| Unknown Genus ↑ |
Note: “↑” means increase in bacteria in diabetic patients vs. n ormal subjects, “↓” means decrease in bacteria in patients vs. normal subjects, superior numbers are related reference numbers. Butyrate-producing bacteria are indicated asterisk (*).
Table 1: Alterations in intestinal microbiome on type 2 diabetic patients.
Second study in China revealed a decreased bacterium of butyrate- producing F. Prausnitzii in prediabetic patients than those showing normal glucose tolerance [66]. As SCFAs, butyrate is a strong energy source for intestinal mucosa and has powerful effects on various gut mucosal functions, working the colonic defense barrier and suppressing oxidative stress [67]. Butyrate was known to enhance the intestinal barrier by regulating the assembly of Tight Junctions (TJs) through the activation of AMP Protein Kinase (AMPK) as well [68]. Decreased Roseburia and Eubacterium have been noted in patients with symptomatic atherosclerosis compared with healthy subjects, working as stenotic atherosclerotic plaques in the area of carotid artery with cerebrovascular events [69]. The counts of the Clostridium coccoides group, Atopobium cluster and Prevotella were decreased, while total Lactobacillus were increased in feces of Japanese type 2 diabetic patients than in those of control subjects [70].
Previous reports have revealed a decline in Akkermansia muciniphila (A. muciniphila) in patients with obesity and diabetes [70]. Zhang et al. reported that the amount of A. muciniphila is relatively low, showing the potential to affect host metabolism deeply [66]. A. muciniphila belongs to the Verrucomicrobia phylum and it colonizes the mucus layer in the gastrointestinal tract, representing 1% to 4% of the fecal microbiota. It stimulates mucosal microbial networks and it improves intestinal barrier function, providing crucial host immunological responses [71]. Zhang et al. further considered that the pasteurized A. muciniphila appeared to be more effective than the live A. muciniphila in HFD-Streptozotocin treated animals [72]. This mucin-degrading bacterium was known to reverse high-fat diet-induced metabolic disturbance: fat-mass gain, metabolic endotoxemia, adipose tissue inflammatory change and insulin resistance as well [73]. A. muciniphila also increases the intestinal contents of endocannabinoids controlling the inflammatory state, gut barrier as well as gut peptide secretion [73]. It has been observed that A. muciniphila can improve glucose metabolism by mediating the negative effects of IFNγ on the host [74]. A. muciniphila exerts a key role in the maintenance of intestinal health and in metabolic modulation [71]. Bu et al. demonstrated that A. muciniphila plays a key role in protecting the expression of tight junction protein as well as intestinal barrier [75]. Overweight and fatty adult patients with increased gene richness and elevated A. muciniphila abundance showed the healthiest metabolic conditions, especially in fasting plasma glucose and triglyceride levels as well as body fat distribution. Subjects with elevated baseline A. muciniphila content showed greater improvement in insulin sensitivity markers and other clinical parameters on calorie restriction for 6-weeks [76]. Many studies further have revealed that F. prausnitzii abundance is decreased in different intestinal disturbances including diabetes type 2 [65,66,77].
In 2015, Lambeth et al. investigated the characteristics of the gut microbiota in prediabetes and type 2 diabetes, compared with non-diabetic subjects [78]. Collinsella and an unknown genus belonging to family Enterobacteriaceae were increased in patients with type 2 diabetes. The increase in Collinsella in type 2 diabetes, noted in the study by Zhang et al. as well has been related to symptomatic atherosclerosis in the study by Karlsson et al. [66,69]. Sato et al. recognized that the fecal levels of total organic acids, acetate and propionate were lower and that the levels of fecal total organic acids were closely related to carbohydrate intake and negatively associated with the uptakes of total fat and saturated fatty acids in patients with type 2 diabetes [70]. Organic acids in feces augment elimination of Escherichia coli O-157 [79]. SCFAs is known to stimulate the release of gut hormones like Glucagon Like Peptide-1 (GLP-1) and Peptide YY (PYY) from intestinal L-cells [80,81]. Conversely, the low fecal concentrations of organic acids may be harmful by inducing disturbances of glycemic control through the reduction of postprandial incretin secretion and the increased susceptibility to infection in type 2 diabetic patients [70].
Extracellular Vesicles (EVs), lipid bilayer structures secreted from the gut microbiome, have recently come into the spotlight because gut microbe derived EVs influence glucose metabolism by enhancing insulin resistance. Nah et al. investigated the co-occurrence of fecal microbes and microbe derived EVs across serum and urine in human subjects, revealing the dynamics and stability of gut derived EVs [82]. The co-occurrence of microbes was compared between patients with Type 2 Diabetes Mellitus (T2DM) and healthy subjects. As a result, higher correlations between stool, serum and urine microbiomes in patients with T2DM compared to healthy subjects indicating the dysfunction of intestinal permeability in T2DM.
Recently there have been several trials of Fecal Microbiota Transplantation (FMT) in fatty patients. FMT from a lean donor in obese metabolically uncompromised patients by oral capsules was found to be safe, well tolerated and led to a sustained microbiome and bile acid profiles similar to that of a lean individual at week 12. Although Allegretti et al. did not find a meaningful change in intestinal hormone Glucagon-Like Peptide 1 (GLP1), they did notice implantation of 9 OTUs in the butyrate-producing and bile-hydrolyzing genus Faecalibacterium which may lead to a decrease in primary bile acids [83]. This reduction in taurocholate may represent a restoration of microbiome Bile Salt Hydrolase (BSH) functionality by FMT.
Gaike et al. reported that the genera Akkermansia and Blautia decreased significantly in treatment-naive diabetics and were restored in patients with type 2 DM on antidiabetic care [84]. Comparative studies on the microbial community in patients of different diabetic states are still lacking. Understanding the transition of microbiome and its relation to serum biomarkers in DM patients with various disease states may open new therapeutic approaches for T2DM. Ding et al. considered in his recent review that the gut microbiota is closely associated with the regulation of the circadian clock in the development of type 2 diabetes, which provides potential for gut microbiota-directed therapies to ameliorate the effects of circadian disruptions linked to the occurrence and development of type 2 diabetes [85].
Several studies have shown a strong correlation between Periodontal Disease (PD), especially in the presence of Porphyromonas gingivalis (Pg) oral infection and systemic disease like type 2 DM [86].The gut microbiota works for the control of energy homeostasis partly through fermentation of dietary fibers producing Short-Chain Fatty Acids (SCFAs), promoting the secretion of the incretin Glucagon-Like Peptide-1 (GLP-1) through binding to the SCFA receptors FFAR2 and FFAR3 on enteroendocrine L-cells [87]. A better understanding of the mechanisms for the regulation of L-cell functions is important to identify new treatment targets to increase the incretin effect through regulation of endogenous synthesis and secretion of GLP-1 [86].
Oral infection by Periodontopathogenic bacteria (Pg) affects gut microbiome dysbiosis and joint destruction via increased Citrullinated Protein (CP) generation in an experimental arthritis model mouse [86]. These authors concluded that Pg administration in SKG mice as the following:
(1) Continued Pg inoculation results in the alterations of gut microbiota.
(2) Dysbiosis of gut microbiota induces inflammation in intestinal tissues.
(3) Increased CP in the intestine enhances systemic Anti-Citrullinated Protein/Peptide Antibody (ACPA) production [86].
Sasaki et al. also emphasized that maintaining excellent oral health is important for systemic health including diabetes mellitus [88].
Although recent studies have indicated that gut microbiome dysbiosis was associated with the onset of T2DM, information on the role of blood microbiome in T2DM development is scarce [89]. The mean diversity of the blood microbiome was not different between T2DM cases and non- T2DM controls. At genus level, the Aquabacterium, Xanthomonas and Pseudonocardia were presented with lower abundance, while Actinotalea, Alishewanella, Microbacterium sediminis and Pseudoclavibacter were presented with higher abundance among T2DM cases compared to those in non-T2DM controls. Participants carried the genus Bacteroides in blood were significantly associated with a decreased risk for T2DM development, with 74% vs. 88%. However, participants carried the genus Microbacterium sediminis have an increased risk for T2DMs [89].
The intestinal barrier protects the entry of pathogenic microorganisms and toxic luminal substances and regulates the absorption of nutrients, electrolytes and water from the intestinal lumen into the circulation [90]. These functions are kept by a complex multilayer system by a mucus layer and epithelial monolayer cells interconnected by TJs. An intact intestinal barrier inhibits the permeation of antigens, pathogens, endotoxins and other pro-inflammatory substances into the human body, whereas intestinal barrier disfunction allows their entry, which might trigger local and systemic inflammation and disease [91]. An anomalous or imbalanced gut microbiota possibly favors an increased intestinal permeability, predisposing to portal translocation of microorganisms, their products and cell wall components [92]. Assessment of gut barrier function in humans is currently possible by using gut permeability assays and by the assessment of biomarkers of epithelial integrity including soluble adhesion molecules, or bacterial markers like endotoxin.
Patients with NAFLD were showed to increase gut permeability caused by disruption of the intercellular TJs characterized by decreased TJ protein ZO-1 expression, which may become the underlying mechanism of translocations of bacteria and their products [22]. Intestinal permeability is augmented in children with NAFLD as well and expresses the grade of steatohepatitis [24]. In 2014, Horton et al. showed that intestinal permeability estimated with Cr-EDTA urinary recovery was augmented in patients with type 2 DM and that it was related to increased values of systemic inflammatory markers representing as high-sensitivity C-reactive protein, IL-6 and TNF-α [93]. Zhang et al. reported that serum zonulin levels, useful markers of intestinal permeability, was elevated in newly diagnosed Chinese patients with Type 2 DM and the zonulin level was related to dyslipidemia, inflammation and insulin resistance [94]. Blood zonulin level was found to be also positively correlated with body mass index (BMI), fasting insulin, triglycerides and IL-6 levels, while it was negatively related to insulin sensitivity in Caucasian male patients [95]. Genetically obese ob/ob and db/db mice revealed increased intestinal permeability, deeply modified distribution of occludin and ZO-1 in the intestinal mucosa and elevated circulating levels of inflammatory cytokines and endotoxin in the portal blood compared with lean control mice [96]. In mice fed high-
fat diet, bacterial products together with complete living bacteria can be translocated from the intestinal lumen into adipose tissues [97].
Gut bacteria-epithelial cell interactions have been suggested as a key contributor of epithelial permeability in several segments of the gastrointestinal tract. In fact, several studies support the notion that bacteria can regulate TJ expression and assembly and thus regulate trans- epithelial permeability [98]. Comprehensive knowledge about the role of the gut microbiota on intestinal barrier function from a highly relevant model, i.e., epithelial organoids, will be crucial in developing strategies to resolve epithelial barrier dysfunctions in several non-infectious chronic diseases [98].
Lipopolysaccharide (LPS), often named as endotoxin, represents the major constituent of the outer cell membrane of gram-negative bacteria and crosses the gut mucosal membrane to enter the blood and directly enhances inflammatory pathways. It can reach the deranged paracellular TJ or can reach the enterocytes together with damaging lipoproteins, because it has a strong affinity to chylomicrons [99].
Both animal experiments and human studies have revealed that a high-fat diet is able to modulate the gut microbiota and increase blood levels of endotoxin [63]. Morbidly fat patients with the highest postprandial hypertriglyceridemia revealed an increase in the endotoxin levels in serum and the chylomicron fraction after eating fatty diet [100]. Although baseline endotoxin level was elevated in patients with type 2 DM and Impaired Glucose Tolerance (IGT) compared with nonobese control subjects, ingestion of a high-fat meal further induced a rise in endotoxin levels in type 2 diabetic, IGT and obese patients [101]. This indicates that a continual snacking enhances their pro-inflammatory conditions in type 2 diabetic and IGT subjects attributable to the constant exposure to circulating endotoxin [101].
Besides paracellular leakage of endotoxin across the intestinal epithelium, there exits more physiological route of endotoxin entry. Namely, the internalization of gram-negative bacteria through TLR4 and myeloid differentiation protein-2 (MD-2)-dependent mechanism [101-104]. Endotoxin is more likely transported to Golgi compartment of the enterocyte, where newly assembled chylomicrons are destined to be located prior to basolateral secretion [105-107]. Chylomicrons are likely to bind endotoxin easily and stimulate endotoxin uptake by enterocyte [107-109]. In fact, the morbidly obese patients with the elevated postprandial hypertriglyceridemia revealed a significant increase in endotoxin levels in the blood and the chylomicron fraction after the fat overload [100]. Although chylomicrons inhibit endotoxin toxicity and cell activation, excess endotoxin attributable to paracellular leakage may strongly enhance inflammatory states in type 2 diabetics [108]. In accordance with these findings, a large cohort study revealed that endotoxemia was tightly associated with cardiometabolic disorders [110].
The lack of endotoxin tolerance in macrophages of type 2 diabetic patients possibly be explained by the fact that low-dose subclinical endotoxemia induces low-grade inflammation via IRAK-1 and Tollip and fails to activate the classical Nuclear Factor-κB (NFκB) pathway causing an anti-inflammatory resolution [111]. Moreover, as described below, obesity may possibly reduce the production of adiponectin [41], which is known to promote endotoxin tolerance [112,113]. Kupffer cells are hepatic macrophages expressing high levels of CD 163. Gut-microbiota alterations and bacterial translocation may attribute to hepatic inflammation. Lipopolysaccharide stimulates toll-like receptor 4 leading to an activation of Kupffer cells which express the surface receptor, CD 163 [114].
Figure 1 summarizes the relationships of gut dysbiosis, intestinal hyperpermeability and endotoxemia in the progression of metabolic syndrome and type 2 diabetes.

Figure 1: Gut dysbiosis, intestinal hyperpermeability and endotoxemia in the grade of metabolic syndrome and type 2 diabetes. Disturbance of Intestinal epithelial barrier and increased permeability together with gut dysbiosis may lead to the translocation of bacterial fragments and endotoxemia, which eventually enhances low-grade inflammation in various tissues and insulin resistance on type 2 diabetes. Inflammatory changes in the adipose tissue of fatty patients show pro-inflammatory cytokinemia. Toll-like receptors induce inflammatory changes and insulin resistance, resulting in an impairment of insulin signaling and β-cell dysfunction. Inflammatory changes in the arterial vessels and liver promotes ischemic heart disease and chronic liver disease, respectively.
Increased intestinal permeability is in this way thought to enhance microbial translocation, metabolic endotoxemia and low-grade inflammatory changes in obese patients with NASH and type 2 diabetes [115]. Low- grade chronic inflammation prevailing in these patients was presumably triggered by activation of TLR4 and TLR2. TLR4 is activated by endotoxin and also by fatty acids, which results in activation of NF-κB and release of pro-inflammatory cytokines such as IL-6, IL-1β, TNF-α and Monocyte Chemotactic Protein-1 (MCP-1) [116].
Human adipose tissue is known as an active site for innate immune response, through activation of TLRs and downstream NF-κB signaling [117]. It also contains many macrophages and, in this way, may function as a first line of defense against superficial various wounds. To deal with evident positive energy balance, the adipocytes in obese patients are destined to develop chronic low-grade inflammation, which provokes secondary effects on other organs including muscle and liver through the inflammatory adipocytokine [117]. The adipose tissue in obese and NASH patients may be enlarged with both hyperplasia and hypertrophy, together with increased macrophage infiltration [118], where adipocytes and recruited macrophages trigger the inflammatory responses via overexpression of TLR2, TLR4 and MyD88 [117,119]. In contrast, anti-diabetic, anti-inflammatory and anti- atherogenic adiponectin expressions are decreased in the adipose tissue of fatty subjects [120]. Their productions are decreased in subjects with visceral fat accumulation and their plasma levels are reversely correlated with visceral adiposity [41]. Obesity-related immune cell infiltration, inflammation and increased oxidative stress enhance metabolic impairments in the insulin-sensitive tissues and as a result, insulin resistance, organ failure and premature aging occur [121]. Hyperglycemia and the following inflammation are the causes of micro- and macro-angiopathies in the circulatory system in diabetic patients. They also promote the gut dysbiosis, increased intestinal permeability and fatty liver disease [121].
Leptin is an adipokine which works to decrees food intake and to increase energy expenditure [122]. Although whole leptin levels are increased in obese patients, its action is not increased due to the condition named leptin resistance [123]. Rajala et al. described that pair-fed leptin receptor-deficient (db/db) mice showed marked alterations in expression of various antimicrobial peptides and a shift in fecal microbiota toward a reduction in the Bacteroidetes to Firmicutes ratio [124]. These imply that the leptin receptor signaling plays a role in modulating microbiota composition, although it is not still clear whether leptin signaling adjust antimicrobial peptides regulating the microbiota, or does leptin signaling directly affect the microbiota [125]. It is reasonable that decreased leptin signaling is likely to enhance SIBO or gut dysbiosis along with unhealthy dietary habit in obese diabetic patients. Clinical data are still lacking to demonstrate the issue at present, but this is worth investigating.
Low-grade inflammatory changes in metabolic syndrome enhances NAFLD in the liver. It enhances a spectrum of pathological changes ranging from the simple fatty liver (NAFL) through NASH to fibrosis and eventually cirrhosis and final hepatocellular carcinoma [44]. Patients with type 2 DM have in this way a twofold to threefold higher risk of dying due to chronic advanced liver diseases associated with a non-virus and non-alcohol- related etiology, largely attributable to NAFLD and NASH [126]. Patients with NASH show endotoxemia and overexpression of TLR4 signaling in the liver associated with pro-inflammatory cytokine release together with systemic inflammation [127,128]. Plasma endotoxin levels and hepatic TLR4 mRNA expression were found to be higher in NASH patients compared with NAFL patients [129]. SIBO often observed in NASH patients is also associated with enhanced hepatic expression of TLR4 and release of IL-8 [46].
Type 2 diabetic patients also show increased TLR2, TLR4 mRNA and proteins in the Peripheral Blood Mononuclear Cells (PBMCs) compared with control subjects [130]. The increased TLR expressions were closely related with Body Mass Index (BMI), Homeostasis Model Assessment-Insulin Resistance (HOMA-IR), glucose, HbA1c and blood free fatty acid levels [130]. Another study revealed that increased TLR4 and TLR2 expressions in PBMCs were correlated with TNF-α and IL-6 expressions in PBMCs and fasting blood glucose and HbA1c levels [119]. The increased circulatory ZO-1 and endotoxin levels were correlated to inflammatory markers and poor glycemic/lipid control [131].
Diabetic complications are related partly with the release of endogenous TLR ligands leading to activation of TLRs signaling [132]. TLR1, 2, 4 and 6 mRNA expressions were augmented evidently in wounds of patients with type 2 DM compared with wounds in non-diabetic subjects [133]. Although several experiments have told that other pattern recognition receptors NOD-1 and TLR9 are related to low-grade inflammation and insulin resistance, so far no study has demonstrated the increases in their ligands, peptidoglycan moieties and bacteria-derived Cytosine Phosphate Guanine (CpG)-containing DNA, respectively, in fatty patients or subjects with type 2 DM [107]. Together with endotoxin, glucose solution itself is known to activate TLR4 expression and to enhance TNF-α and IL-6 secretion in the abdominal subcutaneous adipose tissue and isolated subcutaneous adipocytes [134]. It is demonstrated to induce similar TLRs expression and cytokine secretion of PBMC [135].
Insulin as a chief modulator of glucose homeostasis initiates its physiological effects through activation of the insulin receptor. After tyrosine autophosphorylation, Insulin Receptor Substrate (IRS)-1 and IRS-2 bind and activate Phosphatidylinositol 3-Kinase (PI3-K), increasing serine phosphorylation of Akt, which results in glucose transport in the muscle and adipose tissue, glycogen synthesis in the liver and muscle and lipogenesis in the adipose tissue [107]. Glucose, lipids and endotoxin are chief three factors leading low-grade inflammation and insulin resistance [117]. Lipotoxicity with elevated circulating free fatty acids are related to increased insulin resistance [117]. TLRs, especially TLR4 and TLR2, provoke insulin resistance, which is important in the progression of obesity and metabolic syndrome [116]. Activation of inflammatory pathways due to TLRs enhancement results in a disturbance of the insulin signaling, including decreased phosphorylation of the insulin receptor, IRS and Akt and increased inhibitory serine phosphorylation of IRS-1 [107]. Enhancement of TLR4 by endotoxin in preadipocytes augments the expression of TNF- α and IL-6, which impair the insulin signaling in adipocytes [136]. Endotoxin can promote the expression of iNOS and thus interfere with the insulin signaling [137]. Excessive production of nitric oxide augments insulin resistance by hampering Lipoprotein Lipase (LPL) activity and increasing lipolysis and circulating fatty acids [107,138].
Low-grade inflammation and innate immune system activation further lead to β-cell failure [132]. TLR4 expression is elevated in fat, muscle and pancreatic islet cells, including β-cells and resident macrophages in insulin- resistant mice [139]. Expression of TLR4 in db/db mouse islets increased in parallel with hyperglycemia, which was associated with increased expression and secretion of TNF-α, IL-1 and IL-6 [139]. Endotoxin impairs insulin gene expression (PDX-1 and MafA mRNA levels) of human and rat islets via TLR4 and NF-κB signaling [140]. It is important that the effects of endotoxin on the insulin gene in human islets are observed at concentrations similar to the circulating levels during endotoxemia suggesting that direct repression of the insulin gene may contribute to the metabolic disturbances associated with alterations of the gut microbiota [140].
On the other side, TLR4 or TLR2 deletion improved diet-induced insulin resistance and inflammation of adipose tissue in mice [132]. Diabetic islets have 40% fewer TLR4 positive β-cells, but twice the number of TLR4 positive macrophages as compared to healthy islets [141]. The TLR4 responses elevated in the diabetic mouse islets, which is mainly mediated by newly recruited macrophages [141]. The TLR4 positive macrophages induce apoptosis of β-cells and induce β-cell dysfunction measured as reduced glucose stimulated insulin secretion in mouse islets [141].
Metformin
As a biguanide derivate, metformin has been used widely in the treatment of type 2 diabetes for over 50 years. It gives the major clinical advantage of not inducing hypoglycemia or weight gain and ameliorates hyperglycemia showing remarkable cardiovascular safety [142]. The main effect of this drug is to decrease hepatic glucose production through a transient inhibition of the mitochondrial respiratory-chain complex 1 [143]. The resulting decrease in hepatic energy status activates the AMPK, a cellular metabolic sensor, providing a generally accepted mechanism for metformin effect on hepatic gluconeogenesis [143]. Metformin has pleiotropic effects beyond glucose reduction, including improvement of lipid profiles and lowering microvascular and macrovascular complications associated with type 2 diabetes [144]. These effects have not been totally due to AMPK activation, because intravenous metformin is less effective than oral medication, suggesting important gut pharmacology of the drug [144]. Napolitano et al. found that metformin withdrawal was related to a reduction of GLP-1 and increase in serum bile acids, especially cholic acid and its conjugates [144]. Microbiota abundance of the phylum Firmicutes was positively correlated with changes in cholic acid and its conjugates. On the other side, Bacteroidetes abundance was negatively correlated with them [144]. This means that metformin is considered to enhance GLP-1 secretion and suppress serum bile acids levels and fecal Firmicutes/ Bacteroidetes ratio. Clarembeau et al. performed clinically validate a stable- isotope-dilution LC/MS/MS method for the quantitative analysis of numerous primary and secondary BAs and showed that BAs whose circulating levels are associated with T2DM include numerous 12α-hydroxyl BAs (taurocholic acid, taurodeoxycholic acid, glycodeoxycholic acid, deoxycholic acid and 3-ketodeoxycholic acid), while taurohyodeoxycholic acid was negatively associated with diabetes [145].
Sun et al. found that metformin works through lowering levels of Bacteroides fragilis in the gut, resulting in a decrease in the enzyme bile salt hydrolase and a subsequent increase of hydrophilic Glycoursodeoxycholic Acid (GUDCA) [146]. This may inhibit intestinal FXR and increase the level of liver BAs, leading to improvement of metabolic dysfunction including hyperglycemia [147].
Interestingly, metformin is further known to stimulate mucin formation by the microbiome Akkermansia muciniphila. Metformin is known to induce intestinal mucin 2 and mucin 5 expressions and increases Akkermansia in a culture system [148]. Metformin and Akkermansia administration were also associated with the downregulation of elevated IL-1β and IL-6 mRNA expression in visceral adipose tissue of mice fed a high-fat diet, which suggests that metformin like Akkermansia improves the metabolic profile of diet-induced obesity by ameliorating low-grade tissue inflammation [149]. Oral administration of metformin to mice with LPS-induced endotoxemia or ob/ob mice lowered the plasma and tissue levels of TNF-α and IL-6 and increased Activating Transcription Factor-3 (ATF-3) expression in spleen and lungs, which further supports that metformin exhibits anti-inflammatory action in macrophages [150]. Moreover, high-dose metformin provides anti-inflammatory effects through protecting against oxidative stress and extending the lifespan of middle-aged mice by approximately 6% [151].
Atherosclerosis-associated cardiovascular disease is a chief complication of type 2 DM, where inflammation plays a pivotal role [152]. Metformin has been regarded as a first selected drug for most patients, because it reduces cardiovascular morbidity and mortality [153]. Further human study is needed concerning its pharmacological effects on gut dysbiosis and inflammatory parameters for cardiovascular advantage.
DPP-4 inhibitors
The incretin hormones such as Glucose-dependent Insulinotropic Polypeptide (GIP) and GLP-1 are secreted from enteroendocrine cells in the gut and adjust physiological and homeostatic functions related to glucose control, metabolism and dietary intake [154]. Native GLP-1 has a half-life of less than 2 min as a result of analysis by Dipeptydil-Peptidase-4 (DPP- 4) mainly secreted by endothelial cells and rapid renal clearance of both the intact and degraded GLP-1 molecules [155,156]. DPP-4 inhibitors are recently developed anti-diabetic drugs that can improve glucose metabolism by elevating the concentration and duration of active GLP-1[157,158]. IDPP-4 inhibitors in this way stimulate glucose-dependent insulin secretion and decrease glucagon secretion [7].
DPP-4 thus acts as a multifunctional regulatory protease for cytokines, chemokines and neuropeptides involved in inflammation, immunity and vascular function [159]. Anti-inflammatory effects and anti-diabetes property have opened a new possibility for the use of DPP-4 inhibitors. Makdissi et al. described that one of DPP-4 inhibitors sitagliptin acts as a potent anti-inflammatory effect on patients with type 2 DM [160]. After 12 weeks of sitagliptin intake, the mRNA expressions of TLR4, TLR2, TNF-α, JNK-1, IKKβ and CCR-2 in PBMC of patients with type 2 DM decrease significantly compared with the base-line levels. In vitro examination in human THP- 1 macrophages, sitagliptin induced a significant reduction in pyrin domain containing 3 (NLRP3) inflammasome, TLR4 and IL-1β expression [161]. In apolipoprotein E-deficient diabetic mice fed high-fat diet, another DPP-4 inhibitor alogliptin attenuated diabetes-augmented IL-6 and IL-1β expression in atherosclerotic plaques and inhibited TLR4-enhanced upregulation of IL- 6, IL-1β and other pro-inflammatory cytokines by mononuclear cells [152]. Another DPP-4 inhibitor linagliptin was showed to induce down-regulate the pro-inflammatory marker cyclooxygenase-2 and Macrophage Inflammatory Protein-2 (MIP-2) expression in healing wounds from ob/ob mice [162]. Concerning the effect against NASH, 5-week intake of sitagliptin effectively improved methionine/choline-deficient diet-induced steatohepatitis, where it decreased the expression of Cytochrome P450 2E1 (CYP2E1), 4-Hydroxy- 2-Nonenal (4NHE), fibronectin and α-Smooth Muscle Actin (α-SMA) in the liver and decreased the inflammation of adipose tissue [163]. Recently Somm et al. reported a direct beneficial hepatic action of the GLP-1RA liraglutide, with potential translational relevance for T2D patients affected by NASH [164].
We have presented that sitagliptin markedly supressed liver fibrosis in rats via dectease in activated Hepatic Stellate Cells (HSCs) [165]. These suppressive effects were confirmed to associate with dephosphorylation of ERK1/2, p38 and Smad2/3 in the HSCs. Although direct anti-inflammatory actions of DPP-4 inhibitors have been reported in in vitro studies using aorta tissue [158], mononuclear cells [152], macrophages [166], endothelial cells and adipocytes [167,168], their relation to gut dysbiosis, intestinal permeability or metabolic endotoxemia have not been investigated yet. Mashitani et al. presented that alogliptin for 12 months significantly decreased serum ferritin levels in type 2 diabetics with relatively low HbA1c levels [169]. Although the results are now meaningful in the point that DPP4- inhibitors may be beneficial to prevent the disease progression in patients with NAFLD and type 2 diabetes, the mechanism of decrease in ferritin was not considered in discussion. The effect of DPP4-inhibitors on metabolic endotoxemia may deserve further investigation, because both iron and endotoxin are taken up by Kupffer cells and close correlation has been reported between plasma endotoxin and serum ferritin levels in patients with advanced liver disease [170-172].
SGLT2 inhibitors
Two main sodium-glucose cotransporters, SGLT1 and SGLT2 have been recently introduced. SGLT1 enables the small intestine to absorb glucose and contributes to the reabsorption of glucose filtered by the kidney. Another SGLT, SGLT2 is responsible for reabsorption of most of the glucose filtered by the kidney [173]. Variable SGLT2 inhibitors have been accepted as a new class of treatment for Type 2 diabetic patients [174]. By decreasing renal glucose absorption, these agents are aimed to target hyperglycemia independent of insulin secretion or insulin sensitivity [175]. Based on this unique mechanism of action [175], they are also expected as a safe and effective drug with other agents, including insulin and incretin- based therapies [175].
By the way, there are wide varieties in their selectivity for SGLT2 compared with SGLT1: canagliflozin 160-fold, ipragliflozin 570-fold, dapagliflozin 1,200-fold, luseogliflozin 1,770-fold and empagliflozin 2,700- fold [176]. Although empagliflozin is extremely selective, it is associated with lower rates of all-cause and cardiovascular death and lower risk of hospitalization for heart failure [177]. Heart failure–associated endpoints seemed to account for most of the observed benefits in this study [7]. The marked cardioprotective effect of empagliflozin may not be explained by slight decrease of HbA1c level observed in the study. In experimental studies, intake of empagliflozin not only improved hyperglycemia but also stabilized endothelial function of aortic rings and reduced oxidative stress in aortic vessels of diabetic rats induced by streptozotocin [178]. Another study reported that empagliflozin significantly improved markers of oxidative stress 8-hydroxydeoxyguanosine (8-OHdG) in the kidney of streptozotocin-induced diabetic rats [179]. It inhibited inflammatory and fibrotic gene expression such as MCP-1, intercellular adhesion molecule-1 (ICAM-1). Plasminogen activator ibhibitor-1(PAI-1) and Transforming Growth Factor-β (TGF-β) in the diabetic kidney.
Empagliflozin significantly improved cardiovascular outcomes and shows class III–IV symptoms only 10% in diabetic patients. Empagliflozin treatment of type 2 diabetic patients shows no significant effect on hemodynamic parameters after 1 or 3 days and also after 3 months. It leads to rapid and sustained significant improvement of diastolic function [180]. Renal SGLT1 gene expression was suppressed in the study [179]. Although the authors did not analyze SGLT1 action in the intestine, there was a possibility that empagliflozin does inhibit it, thereby decreasing the rate of intestinal glucose absorption.
SGLT1 is expressed in the small intestine and transports glucose and galactose across the apical membrane in a process driven by a sodium gradient created by Na+/K+-ATPase [174]. Elevated mRNA and protein levels for SGLT1 have been reported in the intestine of obese subjects and type 2 DM [181,182]. Intestinal SGLT1 inhibition suppress and delays the glucose excursion following carbohydrate ingestion and augments GLP-1 and peptide YY secretion [174]. The latter is likely attributed to increased glucose exposure of the colonic microbiome and formation of metabolites, such as L cell secretagogues [174]. An increase in colonic microbial production of SCFAs augments barrier function of the colonic epithelium and possibly inhibits metabolic endotoxemia. Taken together, the above cardioprotective effect of empagliflozin might be explained by its anti- inflammatory actions not only in the kidney but also in the intestine
Although the results of succeeding trials of dual SGLT1/2 inhibitors should be carefully evaluated, the possible effects of SGLT2 inhibitors on intestine, gut dysbiosis and metabolic endotoxemia may deserve further investigation.
Other promising drugs
Finally, Glucagon-Like Peptide-1 Receptor Agonists (GLP-1RAs) pose promising anti-inflammatory, anti-obesogenic and extensive cardiovascular and metabolic benefits and pulmonary protective effects, as well as beneficial impact on gut microbiome composition. It can also preserve healthy gut microbiota (prevent microbiome dysbiosis). Hence, GLP-1RAs turns out to be potential candidates for the treatment of patients, also affected by COVID-19 infection with type 2 diabetes mellitus [183].
Dietary Treatment
Dietary treatment based on latest knowledge on the food-microbiome interaction may improve the effects of diabetic medications. Hiel et al. showed that nutritional intervention targets the gut microbiota – even if they will never solve the problem of obesity and diabetes alone-must are considered in the medical follow-up of obese patients [184]. Changes in fecal microbiota were most pronounced after six weeks of low-calorie formula diet, but they were reverted partially until the end of this study. The gut microbiota phylogenetic diversity increased persistently, however. The abundance of Collinsella, which has previously been associated with atherosclerosis, decreased significantly during this weight loss program [185].
In fact, the Westernization of the Asian diet is known to be responsible for the increase in type 2 diabetes in Asian countries [186]. In fact, sucrose intake and medium-chain fatty acid intake, which are related to characteristics of Western diet [186]. Metabolites produced by the gut microbiota could be pathogenic or beneficial to the host. These metabolites might play crucial roles in host biosynthetic and metabolic networks, as well as various immunological and neurobiological processes. Metabolites are the products and intermediates of cellular metabolism [186]. There is a possibility that reducing sucrose intake could help prevent the onset of type 2 diabetes mellitus through prevention of gut dysbiosis in Japanese individuals [187]. Further a number of scientific studies suggesting a positive impact of ω-3 fatty acids and probiotics (individually and combined) on low-grade inflammation is growing and increasing evidence points towards health-promoting effects from these supplements throughout the life span [188].
We can initiate healthy food preferences from the earliest date, thus optimizing the long-term public health benefits [189]. This meticulous dietary treatment finally suppresses the morbidity of patients with diabetes and related cardiovasclular and hepatic events in risky patients with type 2 diabetes mellitus. Consumption of Mediterranean-style Diets (MD) is considered healthy and associated with the prevention of cardiovascular and metabolic diseases and many other diseases. Such beneficial effects of MD might be attributed to high proportion of fibers, mono- and poly- unsaturated fatty acids, antioxidants and polyphenols [190]. It is becoming evident that the microbial metabolites produced by gut microbiome and dietary interactions plays an important role to regulate host metabolism and physiology. MD feeding enhances abundance of probiotics or beneficial bacteria like Lactobacilli, however, future studies focusing on determining how the increased abundance of Lactobacilli benefit host health will be important to conclude the mechanistic views [190]. Short Chain Fatty Acids (SCFA, particularly butyrate) produced by gut microbes may positively influence insulin sensitivity and insulin secretion, with effects that would prevent T2D [191]. Kesh et al. revealed that T2DM negatively contributes to therapeutic outcomes in pancreatic cancer and they conclude that enrichment of a “tumor protective” gut bacteria as well as enrichment of a “stem-like” population contributed to this phenomenon [192].
Li et al. indicate that IL-37 showed a protective role in the elderly type 2 DM patients and might sensitize them to insulin therapy through suppressing the gut microbiota dysbiosis [193]. Our findings suggested that IL-37 may have clinical potential as a novel therapeutic target in the treatment of type 2 elderly DM patients. Phytochemicals other than caffeine appear to account for most beneficial properties of coffee. As in vegetables or fruits, polyphenols and phenolic acids represent a major portion of phytochemicals in coffee beans [194]. Polyphenols are plant secondary metabolites widely exist in plants and fruits. These substances and their diverse metabolites have profound influence on the diversity and complexity of the intestinal microflora. Various studies have been carried out to understand the response of the gut microbiota with polyphenol administration as well as to identify the key microorganisms involved. Dietary polyphenols and their metabolites contribute to the maintenance of energy homeostasis and gut health through modulation of the gut microbiome, intestinal epithelial cellular function and the mucosal immune system. Although the detailed mechanism by which polyphenols interact with the gut microecology is still not yet well characterized, polyphenols appear to influence energy metabolism and promote weight loss by restructuring the intestinal microecology [195]. The review by Koopen et al. also demonstrates that dietary polyphenols and microecology favorably interact to promote multiple physiological functions on human organism [196].
Lind et al. reported that the Swedish complementary diet contributes to a sustainable environment [197]. A daily supplement of inulin-type fructans induced a moderate, but significant increase in fecal levels of Bifidobacteria, total SCFA, acetic acid and propionic acid in patients with type 2 DM [198].
The close relation between the gut microbiome and diabetes has gradually moved step by step from the initial correlation studies, which proved a strong association, to exploring the causality and potential mechanisms. It is very clear that as science looks to the future as this will be a very promising frontier not only as a biomarker for diabetes, but also as a target for potential therapeutic treatments [199]. The treatment with a low-fat diet, the blood glucose and BMI of patients with T2DM were effectively controlled. However, the changes in intestinal flora were complex [200]. The relative abundances of butyrate-producing bacteria, including Anaerotruncus and Roseburia, were significantly lower in the intestinal tract of patients with T2DM than in healthy individuals. Although the difference in intestinal flora between patients with T2DM and controls was great, the structure of the intestinal flora between diabetic patients and healthy individuals gradually tended to become more similar after 6 months of treatment [200]. Van et al. further strongly supported that plant extracts rich
in polyphenols has a power to improve metabolic disorders associated with obesity and metabolic disorders [201]. Increasing SCFA production could be a valuable strategy in the preventing gastro-intestinal dysfunction, obesity and type 2 DM [202]. Dietary fiber can significantly improve the relative abundance of Bifidobacterium, total SCFAs and glycated hemoglobin. However, dietary fiber did not appear to have a significant effect on fasting blood glucose, HOMA-IR, acetic acid, propionic acid, butyric acid and adverse events [203]. The Very Low-Calorie Ketogenic Diet (VLCKD) appears to be more effective than caloric/energy restriction diets for the treatment of several diseases, such as obesity and diabetes [204-207]. In the Western world, dietary therapy is impoverished of fiber. Dietary fiber intake associates with overall metabolic health (through key pathways that include insulin sensitivity) and a variety of other pathologies that include cardiovascular disease [208-210].
The implications of gut microbiota can be also exemplified in other treatment modalities for type 2 DM. Improvement of metabolic effects of a probiotic VSL#3 on rats fed high-fat diet has been reported based on the increase in SCFA butyrate, which stimulates the release of GLP-1. Enrichment of gut microbiota with Lactobacillus reuteri was known to be increased insulin secretion in glucose-tolerant volunteers. As stated in various studies, gut dysbiosis is likely to play a key modulatory role on the disease progression of type 2 diabetes. Marked technological progress in the studies of gut microbiota has opened a novel area of research field in diabetology. However, we should be aware of its limitations and always try to refine the method. Studies using the direct measurement of microbiota function such as metagenomic, transcriptomic and metabolomic assays (i.e., the metabiome) are needed to determine whether changes in bacterial function rather than composition are related to health and disease. Nevertheless, we can further tune the better microbial composition of patients by a skillful diet therapy to get a maximum effect of each pharmacotherapy. Interestingly enough, some probiotics are reported to increase adiponectin levels and to enhance its receptor AdipoR2 gene expression in experimental animals.
The progress in gut microbiology with emerging analytical technologies has a power to bring about a paradigm shift in the diabetes treatment of tomorrow. There is a great possibility that meticulous management of gut microbiota and intestinal functions may suppress metabolic endotoxemia and inflammation and finally improve the prognosis of patients with diabetes. By all means, lifestyle optimization is essential for all patients with diabetes. Dietary treatment on the basis of latest knowledge on the food-microbiome interaction may improve the effects of diabetic drugs. Hiel et al. shows that nutritional intervention targeting the gut microbiota-even if they will never solve the problem of obesity and diabetes alone must be considered in the medical follow-up of obese. Changes in fecal microbiota were most pronounced after six weeks of low-calorie formula diet, but they were reverted partially until the end of this study. The gut microbiota phylogenetic diversity increased persistently, however. The abundance of Collinsella, which has previously been associated with atherosclerosis, decreased significantly during this weight loss program.
In fact, the Westernization of the Asian diet is known to be responsible for the increase in type 2 diabetes in Asian countries. Sucrose intake and medium-chain fatty acid intake, which are related to characteristics of Western diet sucrose intake and medium-chain fatty acid intake, which are associated with characteristics of Western diet. Metabolites produced by the gut microbiota could be pathogenic or beneficial to the host. These metabolites might play crucial roles in host biosynthetic and metabolic networks, as well as various immunological and neurobiological processes. Metabolites are the products and intermediates of cellular metabolism. There is a possibility that reducing sucrose intake could help prevent the onset of type 2 diabetes mellitus through prevention of gut dysbiosis in Japanese individuals. Further a number of scientific studies suggesting a positive impact of ω-3 fatty acids and probiotics (individually and combined) on low-grade inflammation is growing and increasing evidence points towards health-promoting effects from these supplements throughout the life span.
We can initiate healthy food preferences from the earliest date, thus optimizing the long-term public health benefits. This meticulous dietary treatment finally suppresses the morbidity of diabetic patients and related cardiovasclular and hepatic events in risky patients with type 2 diabetes mellitus. Consumption of MD is considered healthy and associated with the prevention of cardiovascular and metabolic diseases and many other diseases. Such beneficial effects of MD might be attributed to high proportion of fibers, mono- and poly-unsaturated fatty acids, antioxidants and polyphenols. It is becoming evident that the microbial metabolites produced by gut microbiome and dietary interactions plays an important role to regulate host metabolism and physiology. MD feeding enhances abundance of probiotics or beneficial bacteria like Lactobacill,i however, future studies focusing on determining how the increased abundance of Lactobacilli benefit host health will be important to conclude the mechanistic views. Short Chain Fatty Acids (SCFA, particularly butyrate) produced by gut microbes may positively influence insulin sensitivity and insulin secretion, with effects that would prevent T2D. Kesh et al. revealed that T2D negatively contributes to therapeutic outcomes in pancreatic cancer and they conclude that enrichment of a “tumor protective” gut bacteria as well as enrichment of a “stem-like” population contributed to this phenomenon.
Li et al. indicate that IL-37 showed a protective role in the elderly type 2 DM patients and might sensitize them to insulin therapy through suppressing the gut microbiota dysbiosis. Their findings suggested that IL-37 may have clinical potential as a novel therapeutic target in the treatment of type 2 DM in the elderly. Phytochemicals other than caffeine appear to account for most beneficial properties of coffee. As in vegetables or fruits, polyphenols and phenolic acids represent a major portion of phytochemicals in coffee beans. Polyphenols are plant secondary metabolites widely exist in plants and fruits. These substances and their diverse metabolites have profound influence on the diversity and complexity of the intestinal microflora. Various studies have been carried out to understand the response of the gut microbiota with polyphenol administration as well as to identify the key microorganisms involved. Dietary polyphenols and their metabolites contribute to the maintenance of energy homeostasis and gut health through modulation of the gut microbiome, intestinal epithelial cellular function and the mucosal immune system. The impaired immune system together with metabolic imbalance also increases the susceptibility of diabetic patients to several pathogenic agents such as the Severe Acute Respiratory Syndrome coronavirus (COVID-19).
Although the detailed mechanism by which polyphenols interact with the gut microecology is not yet well characterized, polyphenols appear to influence energy metabolism and promote weight loss by restructuring the intestinal microecology. The review by Koopen et al. also demonstrates that dietary polyphenols and microecology favorably interact to promote multiple physiological functions on human organism.
Dietary fiber can significantly improve the relative abundance of Bifidobacterium, total SCFAs and glycated hemoglobin. However, dietary fiber did not appear to have a significant effect on fasting blood glucose, HOMA-IR, acetic acid, propionic acid, butyric acid and adverse events.
Fecal Microbiota Transplantation (FMT) seems to have potential clinical applications in treating diabetic patients with gut dysbiosis. Correction of gut dysbiosis with FMT not only results in mucosal healing in the gut, but also has profound systemic, immunological and metabolic response. GLP-1RAs give to be potential candidates for the treatment of patients, also affected by COVID-19 infection with type 2 diabetes mellitus.
The author has no conflict of interest to declare relevant to the subject of this review article and every statement in it.
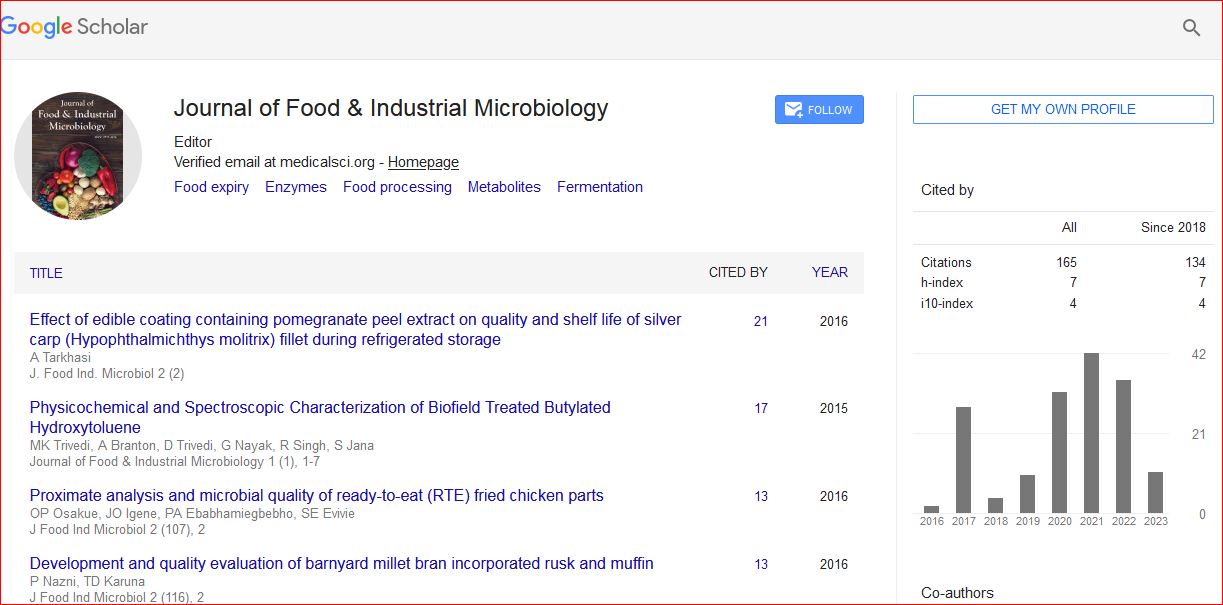
Journal of Food & Industrial Microbiology received 160 citations as per Google Scholar report