Perspective - (2022) Volume 7, Issue 9
Received: 01-Sep-2022, Manuscript No. jidm-22-79847;
Editor assigned: 03-Sep-2022, Pre QC No. P-79847;
Reviewed: 17-Sep-2022, QC No. P-79847;
Revised: 23-Sep-2022, Manuscript No. P-79847;
Published:
01-Oct-2022
, DOI: 10.37421/2576-1420.2022.7.257
Citation: Catalao, Madalena. “The Mycobacterial Cell Envelope is Targeted by Mycobacteriophage Lysis Enzymes.” J Infect Dis Med 7 (2022): 257.
Copyright: © 2022 Catalao M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
At this time, multidrug-resistant bacteria are seen as a danger to world health. This has rekindled interest in using bacteriophages to combat bacterial infections, together with the pharmaceutical industry's disinvestment in the development of novel-acting antibiotics and the urgent need for alternative therapies. This is brought on by phages' bacteriolytic abilities, which cause cell death. By releasing the freshly created phage particles from the infected cells to the environment, double-stranded DNA phages (ds DNA phages) put an end to a lytic cycle. They must produce lysis proteins that exert their action at the proper time in order to go past each barrier of the cell envelope in order to accomplish this. It is this ability of bacteria to lyse that is being investigated for usage. Entire phages or their derived proteins for therapeutic uses, biocontrol in the food industry, and the identification of dangerous bacteria [1].
The intricate structure of the mycobacteria cell membrane distinguishes it from both Gram-positive and Gram-negative bacteria and gives it a special place in the bacterial community. Its extremely high lipid content (up to 60% of the CW is made of lipids) explains the highly hydrophobic cell surface characteristics that result in a natural impermeability to nutrients, resistance to many antibacterial drugs, and exceptional inflammatory activity, playing a key role in virulence. A CM that is physically and functionally similar to that of other bacteria makes up the cell envelope [2]. The CW, which encircles this membrane, is made up of a thin layer of PG covalently connected to an arabinogalactan (AG) polymer, which is then esterified to mycolic acid. acids (MA) composing the mycolyl-arabinogalactan-peptidoglycan complex, which is the CW core (mAGP). The CM and CW are separated by a periplasmic gap. The MAs, one of the distinguishing characteristics of mycobacteria, are long chain fatty acids that are a component of a membrane bilayer known as a genuine OM often referred to as a mycomembrane. Trehalose mono (TMM), di-mycolates (TDM), phospholipids, glycopeptidolipids, and lipoglycans are likely to make up the outer leaflet of this membrane, which is structurally distinct from the OM of Gram-negative bacteria. Trehalose mono (TMM) and di-mycolates (TDM) are significant pathogenic factors in bacteria. Last but not least, a layer called the outermost layer (OL), or the capsule in the case of pathogenic species, is primarily made up of polysaccharides and proteins with a trace quantity of lipids, whereas in nonpathogenic species this OL is made up of lipids consisting largely of proteins [3].
Phage Ms6, a temperate phage that infects the nonpathogenic Mycobacterium smegmatis, has the most thoroughly researched mycobacteriophage lysis system. Five genes, gp1 through gp5, are found in the lysis cassette of Ms6. In a holin-independent manner, gp1 encodes a chaperone-like protein that aids in the endolysin (LysAexport)'s via the host sec system n[4]. The lysis protein (LysB) encoded by the gene gp3 (lysB) is involved in the destruction of the mycobacterium OM and has lipolytic activity. Two holin-like proteins that are encoded by the genes gp4 and gp5 work together to ensure that lysis occurs at the appropriate moment. Gp5, which exhibits properties of class III holins, i.e., a tiny protein with two transmembrane domains, interacts with Gp4, which exhibits properties of a class II holin, with just one transmembrane domain), and it was hypothesised that this complex controls when lysis occurs n [5].
These five genes are not present in every mycobacteriophage lysis cassette, though. Most genomes lack gp1 homologues, and many only have one holin-like gene. While LysA coding genes have been found in all genomes, some genomes lack a holin gene and others lack lysB genes. However, it is obvious that mycobacteriophage-induced lysis is also a three-step process, commencing with holins activity to subvert the CM, which next activates the endolysin function to breakdown the PG meshwork, similar to phages of Gramnegative hosts and because of the presence of an OM. The eradication of the OM is the final phase. The role of LysB is played to break the connection between the OM and AG. The function of the mycobacteriophage endolysins and LysB proteins will be covered in more detail in the next sections as they fall under the purview of this review [6-10].
The current advances in our understanding of the mechanisms of mycobacteriophage-mediated lysis have provided new insights into how mycobacteriophages break down each layer of the mycobacteria envelope and are important for the development of novel tactics to undermine mycobacteria's ability to survive. A highly effective variety of enzymatic activities could be offered by combining the numerous mycobacteriophage endolysin enzymatic activities, which are designed to destroy specific bonds within the mycobacterial PG. This is a tantalising prospect and merits more research. The application of such enzymes to M, an intracellular pathogen, presents an additional barrier. tuberculosis. Recently, it was shown that an endolysin was active against intracellular bacteria. PlyC, an enzyme from a streptococcal phage, has been demonstrated by the authors to be able to pass through epithelial cell membranes and eradicate intracellular Streptococcus pyogenes. Endolysins become increasingly potential for the destruction of pathogenic bacteria as more investigations on them are conducted. The next challenge will be getting the medication to the infection's source. The distribution of antibiotics and other antimicrobial agents, such as endolysins, which demonstrates to be a potent antimicrobial agent, is being investigated by a number of research organisations.
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
Google Scholar, Indexed at, Crossref
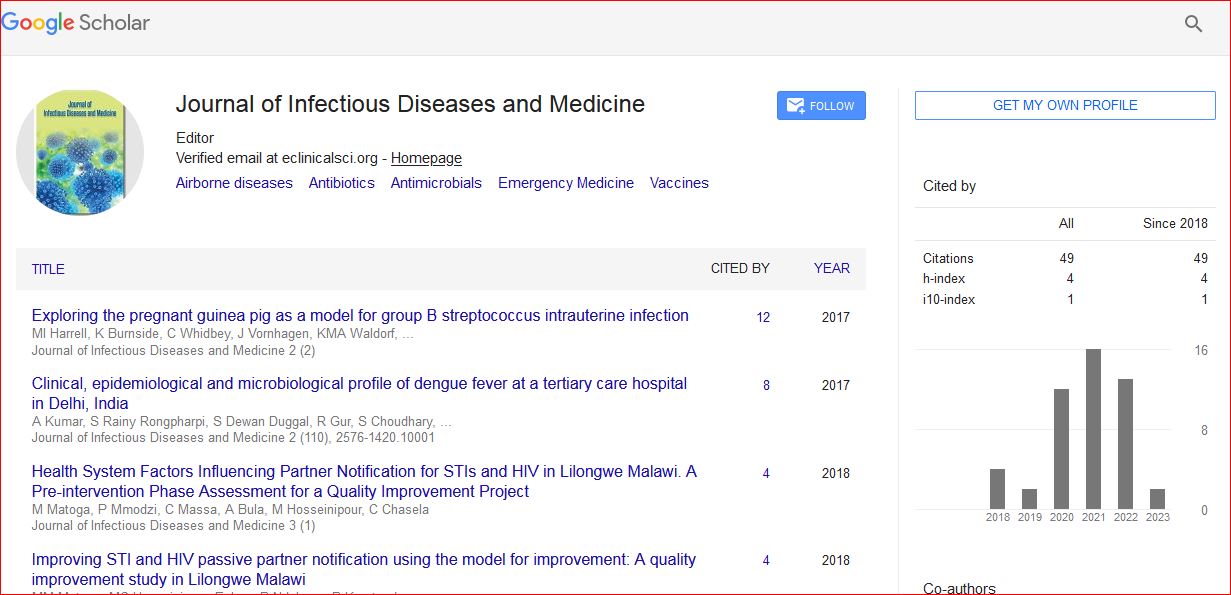
Journal of Infectious Diseases and Medicine received 1059 citations as per Google Scholar report