Perspective - (2022) Volume 7, Issue 3
Received: 08-May-2022, Manuscript No. jib-22-68945;
Editor assigned: 10-May-2022, Pre QC No. P-68945;
Reviewed: 16-May-2022, QC No. Q-68945;
Revised: 24-May-2022, Manuscript No. R-68945;
Published:
29-May-2022
, DOI: 10.37421/2476-1966.2022.7.175
Citation: Horner, Michael. “Viral Hepatitis Immunobiology and Pathogenesis.” J Immuno Biol 7 (2022): 175.
Copyright: © 2022 Horner M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Sources of funding : None
Most cases of viral hepatitis are caused by hepatitis A virus (HAV), hepatitis B virus (HBV), and hepatitis C virus (HCV). Each type of virus infection has a unique clinical outcome and typical natural illness course that are influenced by virological and immunological variables. In contrast to HAV, which does not, HCV frequently leads to a long-lasting infection. HBV is efficiently controlled in adults, despite the fact that it lasts a lifetime following infection in the womb. In this review, we address the immunopathogenesis and immunological responses to HAV, HBV, and HCV infections, which may help to explain the various clinical manifestations and prognoses of each hepatitis virus infection.
Numerous human viruses exhibit hepatotropism, which means that they preferentially infect hepatocytes and result in viral hepatitis, an inflammation of the liver. Hepatitis A virus (HAV), hepatitis B virus (HBV), and hepatitis C virus are responsible for the majority of viral hepatitis cases around the world (HCV). Chronic persistent HCV infection frequently develops after HCV infection. Although HBV can occasionally become a chronic persistent infection, especially when neonates are infected through vertical transmission, acute HBV infection spontaneously cures in more than 90% of infected people. HAV infection leads in an asymptomatic subclinical illness in children but frequently develops in serious liver damage in adults. HAV infection, however, is successfully managed by the host and does not develop into a chronic infection4. Since preventative vaccinations are currently available against HAV and HBV, de novo infection with these viruses is drastically declining in developed nations. A preventative vaccine for HCV is not yet accessible, in contrast [1].
Hepatitis E virus (HEV), a positive-strand non-enveloped RNA virus in addition to these three hepatitis viruses, is a growing public health concern. While HEV often results in an acute, self-limiting infection after faecal-oral transmission, immunocompromised persons can develop chronic illness. In terms of their virological features, HAV, HBV, and HCV vary from one another. The families of Picornaviridae and Flaviviridae, respectively, contain the positive-strand RNA viruses HAV and HCV. The replication of the partly doublestranded DNA virus HBV necessitates reverse transcriptase. Covalently closed circular DNA (cccDNA), a viral transcriptional template and a kind of episomal DNA seen in infected cells, is a major barrier to HBV eradication [2].
Hepatitis E virus (HEV), a positive-strand non-enveloped RNA virus in addition to these three hepatitis viruses, is a growing public health concern. While HEV often results in an acute, self-limiting infection after faecal-oral transmission, immunocompromised persons can develop chronic illness. In terms of their virological features, HAV, HBV, and HCV vary from one another. The families of Picornaviridae and Flaviviridae, respectively, contain the positive-strand RNA viruses HAV and HCV. The replication of the partly doublestranded DNA virus HBV necessitates reverse transcriptase. Covalently closed circular DNA (cccDNA), a viral transcriptional template and a kind of episomal DNA seen in infected cells, is a major barrier to HBV eradication.Direct-acting antivirals (DAAs) can treat chronic HCV infection, but because cccDNA cannot be removed, nucleoside and nucleotide analogue antivirals cannot treat chronic HBV infection [3].
Depending on the virus that causes the infection, HAV, HBV, and HCV infections have typical progressions and consequences. Both the virological properties and the immunological responses that each hepatitis virus elicits dictate the unique courses and results of each infection.
Responses of Type I and type III interferon
The type III IFNs (IFN proteins) and type I interferons (IFN and IFN proteins) are important elements of the innate immune system's antiviral defence. The IFN/ receptor (IFNAR1-IFNAR2) and the IFN receptor (IFNLR1- IL10R2) are the binding sites for type I and type III IFNs, respectively. IFNstimulated gene factor 3 (ISGF3) is a complex made up of phosphorylated signal transducer and activator of transcription 1 (STAT1), phosphorylated STAT2 and IFN regulatory factor 9 that both type I and type III IFNs use to trigger the expression of IFN-stimulated genes (ISGs) (IRF9).
The expression profiles of ISGs in the livers of chimpanzees infected with the virus were studied in IFN responses in hepatitis virus infection. While HAV infection hardly caused ISG expression, HCV infection significantly boosted the expression of many ISGs. Since HBV infection did not result in ISG expression, it was assumed that HBV was a "stealth virus" that the innate immune system was unable to detect [4].
HCV infection
Infection with HCV Retinoic acid-inducible gene I (RIG-I; also referred to as DDX58) and melanoma differentiation-associated protein 5 (MDA5; also referred to as IFIH1) sequentially sense cytosolic HCV RNA. Early on in the infection, the MDA5-mediated IFN response takes over, but the RIGI- mediated IFN response still predominates. EIF2AK2 and Protein Kinase R (PKR) are both involved in the early detection of HCV. PKR interacts with mitochondrial antiviral signalling protein (MAVS) after binding to HCV dsRNA to cause ISGs. By detecting HCV dsRNA in endosomes, toll-like receptor 3 (TLR3) also stimulates the production of IFN and the expression of ISG. Once generated, type I and type III IFNs can inhibit HCV replication through the antiviral activity of ISGs. Pegylated IFN (peg-IFN) has therefore been applied to the management of HCV infection [5].
Natural killer
The natural killer (NK) cells, which are important cellular elements of the innate immune system that fights viruses, perform effector tasks through cytotoxicity and cytokine production that are controlled by the interaction of inhibitory and activating signals. More than ten years ago, a genetic analysis of killer-cell immunoglobulin-like receptors (KIRs) and HLA molecules in HCVexposed individuals showed the significance of NK cells in HCV infection. The natural killer (NK) cells, which are important cellular elements of the innate immune system that fights viruses, perform effector tasks through cytotoxicity and cytokine production that are controlled by the interaction of inhibitory and activating signals.
More than ten years ago, a genetic analysis of killer-cell immunoglobulin like receptors (KIRs) and HLA molecules in HCV-exposed individuals showed the significance of NK cells in HCV infection.
None.
The author shows no conflict of interest towards this manuscript.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
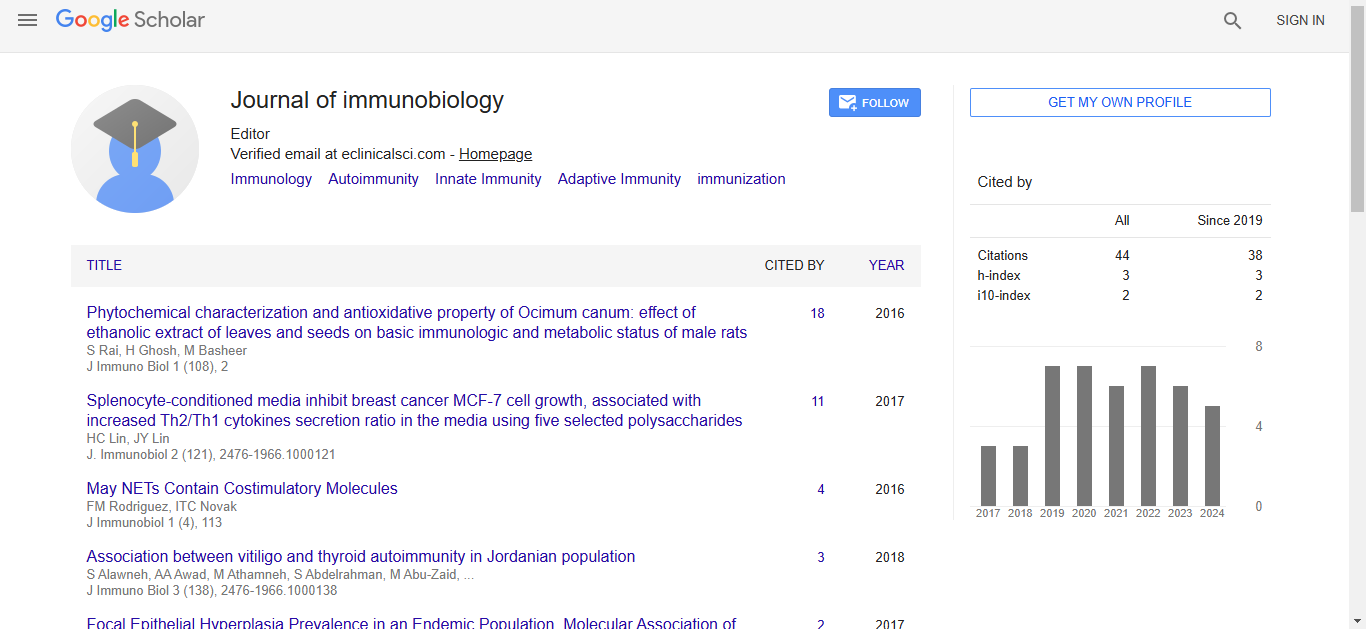
Journal of Immunobiology received 34 citations as per Google Scholar report